Tím chiropraktiky a funkčnej medicíny Back Clinic proti starnutiu. Naše telo je v neustálom a nekončiacom boji o prežitie. Bunky sa rodia, bunky sa ničia. Vedci odhadujú, že každá bunka musí odolať viac ako 10,000 XNUMX individuálnym útokom reaktívnych foriem kyslíka (ROS) alebo voľných radikálov. Bez zlyhania má telo neuveriteľný systém samoliečby, ktorý odoláva útokom a obnovuje to, čo bolo poškodené alebo zničené. Toto je krása nášho dizajnu.
Pochopiť biológiu starnutia a previesť vedecký pohľad na intervencie, ktoré zlepšujú zdravie v neskoršom veku prostredníctvom liečby. Je užitočné mať jasný a konsenzus o tom, čo presne predstavuje ošetrenie proti starnutiu.
Od čias, keď Ponce de Leon hľadal dlhovekosť, človeka vždy lákala šanca na večnú mladosť. Chiropraktická starostlivosť so zdravotným pohybom je silnou metódou stabilizácie a posilnenia tejto samoliečebnej schopnosti. Dr Alex Jimenez diskutuje o konceptoch okolo pandory proti starnutiu.
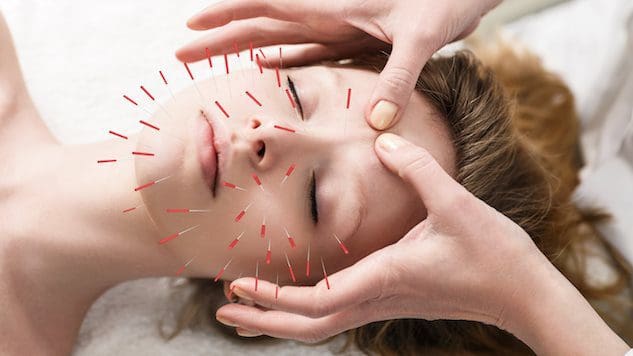
Pre jednotlivcov, ktorí chcú zlepšiť alebo udržať zdravie pokožky, môže začlenenie akupunktúry pomôcť zlepšiť pokožku a bojovať proti procesu starnutia?
Kozmetická akupunktúra
Kozmetická akupunktúra nadväzuje na tradičnú akupunktúrnu prax vpichovania ihly. Cieľom je zvrátiť známky starnutia a zlepšiť zdravie pokožky. Niekedy sa označuje ako akupunktúrne omladenie tváre, ktoré sa používa ako alternatíva k chirurgickým faceliftom a iným konvenčným zákrokom. Predbežné štúdie skúmali, ako môže pomôcť odstrániť starecké škvrny, zdvihnúť ovisnuté viečka a zredukovať vrásky. (Younghee Yun a kol., 2013)
Ako funguje akupunktúra
V tradičnej čínskej medicíne alebo TČM sa akupunktúra už dlho používa na zlepšenie toku energie – čchi alebo čchi – v celom tele. Predpokladá sa, že táto energia cirkuluje cez energetické dráhy známe ako meridiány. Keď sa vyskytnú zdravotné problémy, podľa TČM dochádza k obštrukciám alebo upchatiu obehu.
Akupunkturisti môžu obnoviť optimálnu cirkuláciu / prietok a zlepšiť zdravie vložením ihiel do špecifických akupunktúrnych bodov. (Národný inštitút zdravia, 2007)
Kozmetická akupunktúra
Kozmetická akupunktúra údajne zlepšuje zdravie pokožky a pôsobí ako liečba proti starnutiu tým, že stimuluje produkciu kolagénu. Tento proteín je hlavnou zložkou pokožky. Vnútorná vrstva pokožky starnutím stráca kolagén a pevnosť. Je však potrebný ďalší výskum na podporu tvrdenia, že akupunktúra môže podporovať produkciu kolagénu. Niektorí naznačujú, že kozmetická akupunktúra pomáha omladiť pokožku zlepšením celkovej energie tela. Jedna štúdia zistila, že jednotlivci zaznamenali zlepšenie po piatich sedeniach tvárovej kozmetickej akupunktúry. (Younghee Yun a kol., 2013) Na dosiahnutie optimálnych výsledkov sa však odporúča vykonať desať ošetrení raz alebo dvakrát týždenne. Potom sa udržiavacie kúry vykonávajú každé štyri až osem týždňov. Na rozdiel od botoxu alebo dermálnych výplní nie je kozmetická akupunktúra rýchlym riešením. Dôraz je kladený na vytvorenie dlhodobých zmien na pokožke a tele, čo znamená zlepšenie:
Keď sa ihly vpichujú do kože, vytvárajú rany známe ako pozitívne mikrotraumy. Prirodzené hojivé a opravné schopnosti tela sa aktivujú, keď zacíti tieto rany. Tieto vpichy stimulujú lymfatický a obehový systém, ktorý dodáva kožným bunkám živiny a kyslík a vyživuje ich zvnútra von.
To pomáha vyrovnať pleť a podporuje žiarivosť pleti.
Pozitívne mikrotraumy stimulujú aj tvorbu kolagénu.
To pomáha zlepšiť elasticitu, minimalizovať linky a vrásky.
Alternatívy
Niekoľko prírodných liekov môže pomôcť zlepšiť zdravie pokožky a ponúknuť výhody proti starnutiu. Ceramidy sú molekuly tuku, ktoré sa prirodzene nachádzajú vo vrchnej vrstve pokožky a sú zložkou používanou v produktoch starostlivosti o pleť. Tieto môžu chrániť pred suchosťou pokožky súvisiacou so starnutím. (L Di Marzio 2008) Predbežný výskum naznačuje, že aplikácia bieleho čaju na pokožku môže bojovať proti rozkladu kolagénu a elastínu – proteínu, ktorý podporuje elasticitu pokožky a zabraňuje ochabovaniu). Existujú tiež dôkazy, že prírodné látky, ako je arganový olej, borákový olej a rakytník rešetliakový, môžu ponúkať hydratačné výhody, ktoré by mohli zlepšiť pokožku.(Tamsyn SA Thring a kol., 2009)
Zatiaľ čo sú potrebné ďalšie dôkazy o kozmetickej akupunktúre, integrácia akupunktúry môže pomôcť zvládnuť stres a zlepšiť celkové zdravie. Jednotlivci, ktorí zvažujú kozmetickú akupunktúru, by sa mali poradiť so svojím primárnym poskytovateľom zdravotnej starostlivosti, aby zistili, či je pre nich to pravé.
Spoločné zlepšovanie zdravia: Prijímanie multidisciplinárneho hodnotenia a liečby
Referencie
Yun, Y., Kim, S., Kim, M., Kim, K., Park, JS, & Choi, I. (2013). Účinok kozmetickej akupunktúry tváre na elasticitu tváre: otvorená pilotná štúdia s jedným ramenom. Doplnková a alternatívna medicína založená na dôkazoch: eCAM, 2013, 424313. doi.org/10.1155/2013/424313
Národné centrum doplnkovej a alternatívnej medicíny. (2007). Akupunktúra: Úvod. Webová stránka Národného centra doplnkovej a alternatívnej medicíny. choimd.com/downloads/NIH-info-on-acupuncture.pdf
Kuge, H., Mori, H., Tanaka, TH, & Tsuji, R. (2021). Spoľahlivosť a platnosť hárku na kontrolu tváre (FCS): Kontrolný zoznam pre sebauspokojenie s kozmetickou akupunktúrou. Lieky (Bazilej, Švajčiarsko), 8(4), 18. doi.org/10.3390/medicines8040018
Di Marzio, L., Cinque, B., Cupelli, F., De Simone, C., Cifone, MG, & Giuliani, M. (2008). Zvýšenie hladín kožného ceramidu u starších jedincov po krátkodobej lokálnej aplikácii bakteriálnej sfingomyelinázy zo Streptococcus thermophilus. Medzinárodný časopis imunopatológie a farmakológie, 21(1), 137–143. doi.org/10.1177/039463200802100115
Thring, TS, Hili, P., & Naughton, DP (2009). Antikolagenázové, antielastázové a antioxidačné aktivity extraktov z 21 rastlín. BMC komplementárna a alternatívna medicína, 9, 27. doi.org/10.1186/1472-6882-9-27
Udržiavanie chrbtice jednotlivca v špičkovej forme znamená menej bolesti a väčšiu mobilitu, flexibilitu a slobodu. Telo sa opotrebúva a je prirodzeným prejavom starnutia, ktorý postihuje každého z nás. Problémy s chrbticou súvisiace so starnutím sa môžu stať vážnymi, ak sa neriešia a neriešia cvičeniami, strečingom a chiropraktickou údržbou.
Starnutie a chrbát
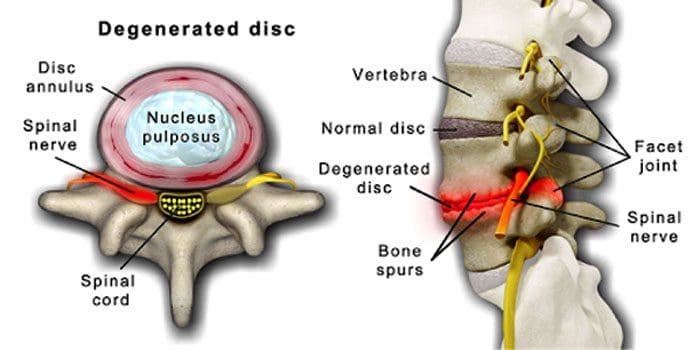
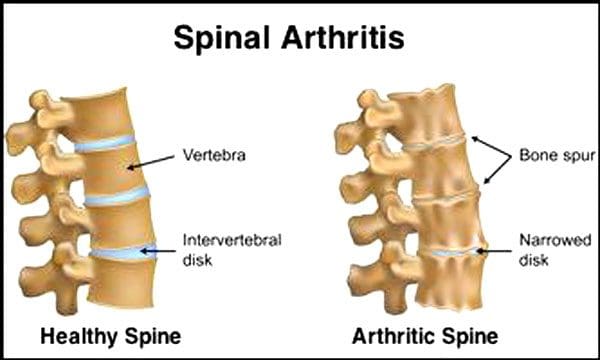
Je normálne, že sa miechové platničky a kĺby vekom zhoršujú. Súčasťou procesu starnutia môže byť aj spinálna stenóza alebo zúženie miechového kanála. Dva stavy spôsobené starnutím sú degeneratívne ochorenie disku a artritída ktoré môžu zahŕňať aj stuhnutie miechových väzov a osteoporóza.
Degeneratívne ochorenie platničiek trpí 40 % jedincov vo veku 40 rokov
Zvyšuje sa na 80 % pre jednotlivcov vo veku 80 rokov a starších.
Sústreďuje sa okolo disky, ktoré sa postupne menia z prevažne vody na prevažne tukové.
Keď je tuk, disky sa zúžia a strácajú elasticitu.
Tvrdia to Centrá pre kontrolu a prevenciu chorôb 23 % dospelých Američanov má artritídu. Ide o stav, ktorý postihuje najmä fazetové kĺby. Kĺby opuchnú, čo znižuje rozsah pohybu a môže naraziť na miechové nervy, čo spôsobuje bolesť, slabosť a ischias. Postupom času väzy okolo a v chrbtici stuhnú, čím sa zníži rozsah pohybu, čo spôsobí stenózu. Strata kostí alebo osteoporóza je spôsobená zmenami hormónov a inými faktormi, ako je výživa. Starnutie je prirodzený proces, ale jednotlivci môžu pomôcť svojej chrbtici zostať v špičkovej forme bez ohľadu na to, koľko majú rokov.
Cvičenie zdravého držania tela
Priamo z voleja správna mechanika zdravého tela je nutnosťou. Zostať si vedomý a mať na pamäti držanie tela udržuje zarovnanie a udržiava telo v rovnováhe. Zdravé držanie tela pomôže znížiť účinky:
Spinálna stenóza
Degeneratívna choroba disku
herniácie
Riziko zlomenín chrbtice
Cvičenie správneho držania tela zahŕňa:
Znížte hrbenie
Uistite sa, že pracovná stanica je v špičkovej forme a ergonomicky zdravá
Pri zdvíhaní dbajte na to, aby ste pokrčili kolená a držte chrbticu čo najvertikálnejšie.
jóga
jóga môže byť veľmi prospešné pre zdravšiu a mladšiu chrbticu. Joga spĺňa tri oblasti pre udržanie chrbtice v top forme, Toto zahŕňa:
Pravidelné cvičenie
Zachováva flexibilitu
Dosahuje ideálnu telesnú hmotnosť
Joga je pre chrbticu aktivita, ktorá bráni veku. Pretože to:
Udržuje pevnosť
flexibilita
Držanie
Zostatok
Môže byť užitočný pri rôznych stavoch chrbtice, najmä pri bolestiach s artritídou
Pády môžu spôsobiť vážne zranenia. Joga môže tiež pomôcť pri práci na rovnováhe.
Navštívte chiropraktika
Preventívna medicína je kľúčom k udržaniu zdravého, mladistvého a čo najpevnejšieho tela. Chiropraktické vyšetrenie môže určiť, či existujú nejaké problémy s chrbticou, a stanoviť diagnózu na vypracovanie optimálneho plánu liečby. Ak je telesná funkcia obmedzená z dôvodu bolesti chrbta a/alebo nôh, kontaktujte Kliniku chiropraktickej a funkčnej medicíny Injury Medical a získajte chrbticu späť v top forme.
Stavba tela
Cvičenie/stabilita Ball Curls
Toto cvičenie pôsobí na svalové skupiny špecifické pre silu chrbtice a zahŕňa:
hamstringy
Lepky
Hlboké brušká
Abduktory a rotátory bedrového kĺbu
Takéto cvičenia sú jedným z najúčinnejších spôsobov, ako vybudovať funkčnú silu a vytrvalosť v oblasti hamstringov, bokov a predchádzať zraneniam. Ak chcete vykonať toto cvičenie:
Ľahnite si na chrbát s pokrčenými kolenami
Zdvihnite nohy tak, aby spodná časť chodidiel spočívala na cvičebnej lopte
Vyvaľujte nohy, kým nebudú rovné
Držte pozíciu na sekundu alebo dve
Vráťte sa na vrchol pohybu a zároveň stláčajte hamstringy
Práca s týmito svalmi vám uľahčí drep, výpady alebo ohýbanie chrbtice.
Zrieknutie sa zodpovednosti za príspevok na blogu Dr. Alexa Jimeneza
Rozsah našich informácií je obmedzený na chiropraktické, muskuloskeletálne, fyzické lieky, wellness a citlivé zdravotné problémy a/alebo články, témy a diskusie o funkčnej medicíne. Funkčné zdravotné a wellness protokoly používame na liečbu a podporu starostlivosti o zranenia alebo poruchy pohybového aparátu. Naše príspevky, témy, predmety a postrehy pokrývajú klinické záležitosti, problémy a témy, ktoré priamo alebo nepriamo podporujú náš klinický rozsah praxe.*
Naša kancelária sa primerane pokúsila poskytnúť podporné citácie a identifikovala relevantnú výskumnú štúdiu alebo štúdie podporujúce naše príspevky. Kópie podporných výskumných štúdií tiež sprístupňujeme rade a/alebo verejnosti na požiadanie. Chápeme, že pokrývame záležitosti, ktoré si vyžadujú dodatočné vysvetlenie, ako to môže pomôcť pri konkrétnom pláne starostlivosti alebo liečebnom protokole; preto, ak chcete ďalej diskutovať o predmete vyššie, neváhajte sa opýtať Dr. Alexa Jimeneza alebo nás kontaktujte na čísle 915-850-0900. Poskytovatelia s licenciou v Texase a Novom Mexiku*
Potraviny, ktoré jeme, môžu mať potenciál byť prospešné alebo škodlivé pre naše zdravie. Zlá výživa môže spôsobiť rôzne zdravotné problémy, vrátane obezity, kardiovaskulárnych ochorení a cukrovky 2. Medzitým vám správna výživa môže spôsobiť, že sa budete cítiť nabití energiou, znížite riziko zdravotných problémov, ako aj pomôžete udržať a regulovať zdravú váhu. Ak chcete podporiť dlhovekosť, musíte svoje telo zásobiť dobrými potravinami. V nasledujúcom článku uvedieme niekoľko dobrých potravín, ktoré môžu v konečnom dôsledku prispieť k dlhovekosti tým, že tiež pomôžu zlepšiť celkové zdravie a pohodu.
Kruhovitá zelenina
Krížová zelenina má jedinečnú schopnosť meniť naše hormóny, spúšťať prirodzený detoxikačný systém tela a dokonca znižovať rast rakovinových buniek. Tie sa musia dôkladne požuť alebo zjesť nastrúhané, nasekané, odšťavené alebo rozmixované, aby sa uvoľnili ich prospešné vlastnosti. Zistilo sa tiež, že sulforafan, ktorý sa nachádza v krížovej zelenine, pomáha chrániť stenu krvných ciev pred zápalom, ktorý môže spôsobiť srdcové choroby. Krížová zelenina, ako je kel, kapusta, ružičkový kel, karfiol a brokolica, sú niektoré z potravín s najvyšším množstvom živín na svete.
Zelený šalát
Surová listová zelenina má menej ako 100 kalórií na libru, čo z nej robí ideálne jedlo na chudnutie. Konzumácia väčšieho množstva zeleného šalátu sa tiež spája so zníženým rizikom srdcového infarktu, mŕtvice, cukrovky a niekoľkých typov rakoviny. Surová listová zelenina je tiež bohatá na esenciálny vitamín B-folát plus luteín a zeaxantín, karotenoidy, ktoré môžu pomôcť chrániť oči. Fytochemikálie rozpustné v tukoch, ako sú karotenoidy, ktoré sa nachádzajú v zeleninových šalátoch, ako je šalát, špenát, kel, kapusta a horčica, majú tiež antioxidačné a protizápalové účinky v tele.
Orechy
Orechy sú nízkoglykemickou potravinou a skvelým zdrojom zdravých tukov, rastlinných bielkovín, vlákniny, antioxidantov, fytosterolov a minerálov, čo tiež pomáha znižovať glykemickú nálož celého jedla, čo z nich robí nevyhnutnú súčasť liečby cukrovky. diéta. Bez ohľadu na ich kalorickú hustotu môže jedenie orechov pomôcť pri chudnutí. Orechy môžu tiež znížiť cholesterol a pomôcť znížiť riziko srdcových ochorení.
Semená
Semená, podobne ako orechy, tiež poskytujú zdravé tuky, antioxidanty a minerály, majú však viac bielkovín a sú bohaté na stopové minerály. Chia, ľanové a konopné semienka sú bohaté na omega-3 tuky. Chia, ľanové a sezamové semienka sú tiež bohaté na lignany alebo fytoestrogény bojujúce proti rakovine prsníka. Sezamové semienka sú navyše bohaté na vápnik a vitamín E a tekvicové na zinok.
Bobule
Bobule sú ovocie bohaté na antioxidanty, ktoré môžu pomôcť pri podpore zdravia srdca. Výskumné štúdie, v ktorých účastníci jedli jahody alebo čučoriedky denne po dobu niekoľkých týždňov, zaznamenali zlepšenie krvného tlaku, celkového a LDL cholesterolu a dokonca aj známky oxidačného stresu. Bobule majú tiež protirakovinové vlastnosti a ukázalo sa, že pomáhajú predchádzať kognitívnemu poklesu spojenému so starnutím.
Granátové jablko
Najznámejšia fytochemikália v granátových jablkách, punicalagin, je zodpovedná za viac ako polovicu antioxidačnej aktivity ovocia. Fytochemikálie z granátového jablka majú protirakovinové, kardioprotektívne a prospešné účinky na mozog. V jednej výskumnej štúdii mali starší dospelí, ktorí pili šťavu z granátového jablka denne počas 28 dní, lepší výkon v pamäťovom teste v porovnaní s tými, ktorí pili placebo nápoj.
Fazuľa
Jedenie fazule a iných strukovín môže pomôcť vyrovnať hladinu cukru v krvi, znížiť chuť do jedla a chrániť pred rakovinou hrubého čreva. Fazuľa je potravinou proti cukrovke, ktorá môže pomôcť pri chudnutí, pretože sa trávi pomaly, čo spomaľuje zvýšenie hladiny cukru v krvi po jedle a pomáha predchádzať chuti do jedla podporou sýtosti. Zistilo sa, že konzumácia fazule a iných strukovín dvakrát týždenne znižuje riziko rakoviny hrubého čreva. Konzumácia fazule a iných strukovín, ako je červená fazuľa, čierna fazuľa, cícer, šošovica a hrach, tiež poskytuje významnú ochranu pred inými druhmi rakoviny.
Huby
Pravidelná konzumácia húb je spojená so zníženým rizikom rakoviny prsníka. Biele huby a huby Portobello sú obzvlášť prospešné proti rakovine prsníka, pretože obsahujú inhibítory aromatázy alebo zlúčeniny, ktoré inhibujú produkciu estrogénu. Ukázalo sa, že huby majú protizápalové účinky a tiež poskytujú zvýšenú aktivitu imunitných buniek, zabraňujú poškodeniu DNA, spomaľujú rast rakovinových buniek a inhibujú angiogenézu. Huby by sa mali vždy variť, pretože surové huby majú potenciálne karcinogénnu chemikáliu známu ako agaritín, ktorá sa varením výrazne znižuje.
Cibuľa a cesnak
Cibuľa a cesnak poskytujú výhody kardiovaskulárnemu a imunitnému systému, ako aj antidiabetické a protirakovinové účinky. Tie sa tiež spájajú s nižším rizikom rakoviny žalúdka a prostaty. Cibuľa a cesnak sú známe svojimi organosírovými zlúčeninami, ktoré pomáhajú predchádzať rozvoju rakoviny detoxikáciou karcinogénov, znižovaním rastu rakovinových buniek a blokovaním angiogenézy. Cibuľa a cesnak majú tiež vysoké koncentrácie zdraviu prospešných flavonoidových antioxidantov, ktoré majú protizápalové účinky, ktoré môžu pomôcť pri prevencii rakoviny.
paradajky
Paradajky sú bohaté na rôzne živiny, ako je lykopén, vitamín C a E, betakarotén a flavonolové antioxidanty. Lykopén môže pomôcť chrániť pred rakovinou prostaty, poškodením kože UV žiarením a? srdcovo-cievne ochorenie. Lykopén sa lepšie vstrebáva, keď sú paradajky varené. Jedna šálka paradajkovej omáčky má asi 10-krát väčšie množstvo lykopénu ako šálka surových nakrájaných paradajok. Majte tiež na pamäti, že karotenoidy, ako je lykopén, sa najlepšie vstrebávajú, keď sú sprevádzané zdravými tukmi, takže si vychutnajte paradajky v šaláte s orechmi alebo v dresingu na báze orechov pre extra nutričné výhody.
Potraviny, ktoré jeme, môžu mať potenciál byť prospešné alebo škodlivé pre naše zdravie. Zlá výživa môže spôsobiť rôzne zdravotné problémy, vrátane obezity, kardiovaskulárnych ochorení a cukrovky 2. Medzitým vám správna výživa môže spôsobiť, že sa budete cítiť nabití energiou, znížite riziko zdravotných problémov, ako aj pomôžete udržať a regulovať zdravú váhu. Ak chcete podporiť dlhovekosť, musíte svoje telo zásobiť dobrými potravinami. Dobré jedlo môže tiež pomôcť znížiť zápal spojený s rôznymi zdravotnými problémami, vrátane bolesti kĺbov a artritídy. Zdravotnícki odborníci, ako sú chiropraktici, môžu ponúknuť rady týkajúce sa stravovania a životného štýlu, aby pomohli podporiť zdravie a pohodu. V nasledujúcom článku uvedieme niekoľko dobrých potravín, ktoré môžu v konečnom dôsledku prispieť k dlhovekosti. – Dr. Alex Jimenez DC, CCST Insight
Šťava z cukrovej repy
Dávky: 1 Doba varenia: 5-10 minút
� 1 grapefruit, olúpaný a nakrájaný na plátky
� 1 jablko, umyté a nakrájané
� 1 celá repa a listy, ak ich máte, umyté a nakrájané
� 1-palcový gombík zázvoru, opláchnutý, olúpaný a nasekaný
Všetky ingrediencie odšťavte v kvalitnom odšťavovači. Najlepšie ihneď podávať.
Len jedna mrkva vám dodá celý denný príjem vitamínu A
Áno, zjedením len jednej uvarenej 80 g (2 �oz) mrkvy získate dostatok beta karoténu, aby vaše telo vyprodukovalo 1,480 900 mikrogramov (mcg) vitamínu A (potrebného na obnovu kožných buniek). To je viac ako odporúčaný denný príjem vitamínu A v USA, ktorý je asi XNUMX mcg. Najlepšie je jesť mrkvu varenú, pretože to zmäkčuje bunkové steny a umožňuje absorbovať viac beta karoténu. Pridanie zdravších potravín do vašej stravy je skvelý spôsob, ako zlepšiť svoje celkové zdravie.
Rozsah našich informácií je obmedzený na chiropraktické, muskuloskeletálne, fyzické lieky, wellness a citlivé zdravotné problémy a/alebo články, témy a diskusie o funkčnej medicíne. Funkčné zdravotné a wellness protokoly používame na liečbu a podporu starostlivosti o zranenia alebo poruchy pohybového aparátu. Naše príspevky, témy, predmety a postrehy pokrývajú klinické záležitosti, problémy a témy, ktoré súvisia a podporujú priamo alebo nepriamo náš klinický rozsah praxe.* Naša kancelária sa primerane pokúsila poskytnúť podporné citácie a identifikovala relevantnú výskumnú štúdiu alebo štúdie podporujúce naše príspevky. Kópie podporných výskumných štúdií tiež sprístupňujeme rade a/alebo verejnosti na požiadanie. Chápeme, že pokrývame záležitosti, ktoré si vyžadujú dodatočné vysvetlenie, ako to môže pomôcť pri konkrétnom pláne starostlivosti alebo liečebnom protokole; preto, ak chcete ďalej prediskutovať vyššie uvedenú tému, neváhajte sa opýtať Dr. Alexa Jimeneza alebo nás kontaktujte na adrese 915-850-0900. Poskytovatelia s licenciou v Texase* a Novom Mexiku*�
Kurátorom je Dr. Alex Jimenez DC, CCST
Referencie:
Joel Fuhrman, MD. �10 najlepších jedál, ktoré môžete jesť, aby ste žili dlhšie a zostali zdraví.� Veľmi dobré zdravie, 6. júna 2020, www.verywellhealth.com/best-foods-for-longevity-4005852.
Dowden, Angela. �Káva je ovocie a ďalšie neuveriteľne pravdivé fakty o jedle.� Životný štýl MSN4. júna 2020, www.msn.com/en-us/foodanddrink/did-you-know/coffee-is-a-fruit-and-other-unbelievably-true-food-facts/ss-BB152Q5q?li=BBnb7Kz&ocid =mailsignout#image=24.
Ak máte niektorú z týchto situácií, potom môžu byť vaše kolagénové peptidy nízke.
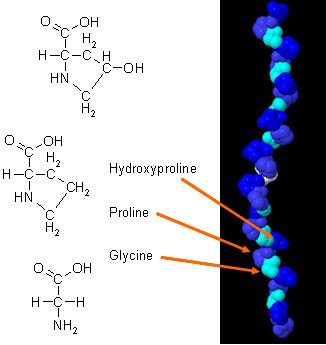
Tam majú boli nové štúdie o tom, ako môže kolagén zlepšiť stavbu tela, keď sa skombinuje s každodenným cvičením. Kolagén v tele má jedinečné zloženie aminokyselín, ktoré hrá zásadnú úlohu v anatómii tela. Kolagénový proteín je koncentrovaným zdrojom glycínu, prolínu a hydroxyprolínu, a keď ho porovnávame so všetkými ostatnými diétnymi proteínmi, robí z kolagénu potenciálnu praktickú voľbu ako štrukturálny proteín.
In Štúdia 2015, výskumníci preukázali, ako môžu účinné doplnky kolagénu zlepšiť zloženie tela u aktívnych mužov. Výsledky ukazujú, že každý muž sa zúčastňuje silového tréningu aspoň trikrát týždenne a musí doplniť aspoň 15 gramov kolagénových peptidov, aby dosiahol maximálne zdravie. Hodnotenia, ktoré tento test poskytuje, sú silový test, bioimpedančná analýza (BIA) a svalové biopsie. Tieto testy zaisťujú, že mužskí jedinci majú dobrý výkon po užití kolagénových doplnkov a výsledky ukazujú, ako sa ich telesná hmotnosť zvýšila bez tuku. Ďalšia štúdia ukázala, ako suplementácia kolagénových bielkovín v kombinácii s odporovým tréningom môže zvýšiť svalovú hmotu a svalovú silu u starších ľudí, ako aj u ľudí so sarkopéniou.
Priaznivé vlastnosti s kolagénom
Existujú veľa prospešných vlastností ktoré kolagénové doplnky môžu telu poskytnúť pri jeho konzumácii. Existujú hydrolyzovaný kolagén a želatína a môžu pomôcť zlepšiť štruktúru pokožky človeka. Aj keď nie je veľa štúdií o kolagénových doplnkoch, existujú skvelé prísľuby pre oblasti na tele. Oni sú:
Svalová hmota: Kolagénové doplnky v kombinácii so silovým tréningom môžu zvýšiť svalovú hmotu a silu v tele.
Artritída: Kolagénové doplnky môžu pomôcť ľuďom s osteoartrózou. Štúdie ukazujú že keď ľudia s osteoartrózou užívajú kolagénové doplnky, zistili masívny ústup bolesti, ktorú zažívali.
Elasticita pokožky: V Štúdia 2014uvádza, že ženy, ktoré užívali doplnky kolagénu a preukázali zlepšenie elasticity pokožky. Kolagén možno použiť aj pri lokálnych ošetreniach, aby pomohol zlepšiť vzhľad pokožky tým, že minimalizuje jemné linky a vrásky.
Nielen kolagénové doplnky poskytujú prospešné vlastnosti špecifickým oblastiam tela, ale existujú štyri hlavné typy kolagénu a aké sú ich úlohy v ľudskom tele, ako aj ich funkcie:
Zadajte 1: Kolagén typu 1 obsahoval 90 % kolagénu v tele a pozostával z husto zložených vlákien, ktoré poskytujú štruktúru pokožke, kostiam, spojivovým tkanivám a zubom v tele.
Zadajte 2: Kolagén typu 2 sa skladá z voľne uložených vlákien, ktoré sa nachádzajú v elastickej chrupavke, ktorá pomáha tlmiť kĺby v tele.
Zadajte 3: Kolagén typu 3 pomáha podporovať štruktúru svalov, orgánov a tepien, ktoré zabezpečujú správne fungovanie tela.
Zadajte 4: Kolagén typu 4 sa nachádza vo vrstvách pokožky každého človeka a pomáha s filtráciou v tele.
Keďže tieto štyri typy kolagénu sú v tele, je dôležité vedieť, že kolagén sa môže časom s vekom prirodzene znižovať, pretože telo produkuje menej kolagénu nižšej kvality. Jedným z viditeľných prejavov úbytku kolagénu je, keď sa pokožka na ľudskom tele stáva menej pevnou a pružnou, ako aj oslabením chrupavky v dôsledku starnutia.
Faktory, ktoré môžu poškodiť kolagén
Aj keď sa kolagén môže s vekom prirodzene znižovať, mnohé faktory môžu zničiť kolagény, ktoré sú pre pokožku škodlivé. Škodlivé faktory môžu zahŕňať:
Cukor a sacharidy: Rafinované cukry a sacharidy môže zasahovať so schopnosťou kolagénu opraviť sa na pokožke. Takže minimalizovaním spotreby cukru a sacharidov v tele môže znížiť účinky dysfunkcie ciev, obličiek a kožného tkaniva.
Expozícia slnku: Aj keď dostatok slnka môže človeku pomôcť užiť si deň, ak je na slnku vystavený dlhší čas môže spôsobiť poškodenie na pokožku a ničí kolagénové peptidy. Následky nadmerného vystavovania sa slnku môžu spôsobiť starnutie pokožky a spôsobiť oxidačný stres v tele.
fajčenie: Keď človek fajčí, môže znížiť produkciu kolagénu v tele, čo spôsobuje, že telo má predčasné vrásky, a ak je telo zranené, proces hojenia bude pomalší a môže viesť k chorobám v tele.
Autoimunitné choroby: Niektoré autoimunitné ochorenia môžu tiež poškodiť produkciu kolagénu, ako je lupus.
záver
Kolagén je pre telo životne dôležitý, pretože pomáha pokožke byť jemná a pevná. Prirodzene, s pribúdajúcim vekom sa bude znižovať, takže užívanie kolagénových doplnkov môže zabezpečiť správne fungovanie tela. Keď na telo pôsobia škodlivé faktory, môžu zastaviť alebo dokonca poškodiť tvorbu kolagénu a urýchliť proces tvorby predčasných vrások, vďaka čomu človek vyzerá staršie ako je. Niektorí Produkty môže pomôcť bunkovej aktivite tela poskytnutím vynikajúcej stability, biologickej dostupnosti a komfortu trávenia.
Rozsah našich informácií je obmedzený na chiropraktické, muskuloskeletálne a nervové zdravotné problémy alebo články, témy a diskusie o funkčnej medicíne. Funkčné zdravotné protokoly využívame na liečbu úrazov alebo porúch pohybového aparátu. Naša kancelária sa primerane pokúsila poskytnúť podporné citácie a identifikovala relevantnú výskumnú štúdiu alebo štúdie podporujúce naše príspevky. Kópie podporných výskumných štúdií tiež sprístupňujeme predstavenstvu a/alebo verejnosti na požiadanie. Ak chcete ďalej prediskutovať vyššie uvedenú tému, neváhajte sa opýtať Dr. Alexa Jimeneza alebo nás kontaktujte na adrese 915-850-0900.
Referencie:
Bosch, Ricardo a kol. �Mechanizmy fotostarnutia a kožnej fotokarcinogenézy a fotoprotektívne stratégie s fytochemikáliami.� Antioxidanty (Bazilej, Švajčiarsko), MDPI, 26. marec 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4665475/.
Danby, F. William. �Výživa a starnutie pokožky: Cukor a glykácia.� Dermatologické kliniky, Národná lekárska knižnica USA, 2010, www.ncbi.nlm.nih.gov/pubmed/20620757.
Jennings, Kerri-Ann. � Kolagén – čo to je a na čo je dobrý?� Healthline, 9. september 2016, www.healthline.com/nutrition/collagen.
Jurgelewicz, Michael. �Nová štúdia demonštruje výhody kolagénových peptidov na zlepšenie stavby tela v kombinácii s cvičením.� Dizajn pre zdravie, 31. mája 2019, blog.designsforhealth.com/node/1031.
Knuutinen, A, a kol. �Fajčenie ovplyvňuje syntézu kolagénu a premenu extracelulárnej matrice v ľudskej koži.� British Journal of Dermatology, Národná lekárska knižnica USA, apríl 2002, www.ncbi.nlm.nih.gov/pubmed/11966688.
Proksch, E, a kol. �Perorálna suplementácia špecifických kolagénových peptidov má priaznivé účinky na fyziológiu ľudskej kože: dvojito zaslepená, placebom kontrolovaná štúdia.� Farmakológia a fyziológia kože, Národná lekárska knižnica USA, 2014, www.ncbi.nlm.nih.gov/pubmed/23949208.
Schauss, Alexander G, a kol. �Účinok nového nízkomolekulárneho hydrolyzovaného výťažku z kuracej hrudnej chrupavky, kolagénu BioCell, na zlepšenie symptómov súvisiacich s osteoartritídou: Randomizovaná, dvojito zaslepená, placebom kontrolovaná štúdia.� Časopis poľnohospodárskej a potravinárskej chémie, Národná lekárska knižnica USA, 25. apríla 2012, www.ncbi.nlm.nih.gov/pubmed/22486722.
Zdzieblik, Denise a kol. �Suplementácia kolagénovým peptidom v kombinácii s odporovým tréningom zlepšuje telesné zloženie a zvyšuje svalovú silu u starších sarkopenických mužov: Randomizovaná kontrolovaná štúdia.� British Journal of Nutrition, Cambridge University Press, 28. októbra 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4594048/.
Informovaním jednotlivcov o tom, ako Národná univerzita zdravotníckych vied poskytuje poznatky budúcim generáciám, univerzita ponúka širokú škálu lekárskych profesií pre funkčnú medicínu.
Ako keby vám bola diagnostikovaná celiakia, syndróm dráždivého čreva, divertikulóza/divertikulitída alebo syndróm netesného čreva?
Nadmerné grganie, grganie, príp nadúvanie?
Abnormálna distenzia po určitých probiotikách alebo prírodných doplnkoch výživy?
Podozrenie na nutričnú malabsorpciu?
Ustupujú tráviace ťažkosti relaxáciou?
Ak zažívate niektorú z týchto situácií, možno máte črevné problémy a možno budete musieť vyskúšať program 4R.
Precitlivenosť na potraviny, reumatoidná artritída a úzkosť sú spojené so zhoršenou gastrointestinálnou permeabilitou. Tieto rôzne stavy sa môžu vyskytnúť v dôsledku mnohých faktorov, ktoré môžu ovplyvniť tráviaci trakt. Ak sa nelieči, môže to byť potenciálne dôsledok dysfunkcie bariéry črevnej permeability, čo môže spôsobiť zápal a vážne zdravotné stavy, ktoré sa môžu vyvinúť v čreve. Program 4R sa používa na obnovenie zdravých čriev v tele a zahŕňa štyri kroky. Sú to: odstránenie, výmena, preočkovanie a oprava.
Črevná priepustnosť
Črevná priepustnosť pomáha chrániť telo a zabezpečuje, aby sa škodlivé baktérie nedostali do čreva. Chráni telo pred potenciálne environmentálne faktory ktoré môžu byť škodlivé a dostávajú sa cez tráviaci trakt. Môže to byť buď toxín, patogénne mikroorganizmy a iné antigény, ktoré môžu poškodiť tráviaci trakt a spôsobiť problémy. Črevná výstelka pozostáva z vrstvy epitelových buniek, ktoré sú oddelené tesnými spojmi. V zdravom črevetesné spojenie reguluje črevnú permeabilitu tým, že selektívne umožňuje látkam prenikať cez črevnú bariéru a prechádzať cez ňu a bráni absorpcii škodlivých faktorov.
Určité environmentálne faktory môžu poškodiť tesné spojenie a výsledkom je, že môže zvýšiť črevnú priepustnosť, čo spôsobuje črevnú hyperpermeabilitu alebo netesné črevá v tele. Prispievajúce faktory môžu zvýšiť priepustnosť čriev, ako je nadmerné množstvo nasýtených tukov a alkoholu, nedostatok živín, chronický stres a infekčné choroby.
So zvýšenou priepustnosťou čriev v čreve môže umožniť, aby antigény prešli sliznicou čreva a dostali sa do krvného obehu, čo spôsobuje imunitnú odpoveď a zápal v tele. Existujú určité gastrointestinálne stavy, ktoré sú spojené s črevnou hyperpermeabilitou a ak sa neliečia, môžu vyvolať určité autoimunitné stavy, ktoré môžu poškodiť telo.
Program 4Rs
4Rs je program, ktorý zdravotnícki pracovníci radia svojim pacientom, aby ho používali, keď riešia rušivé tráviace problémy a pomáha podporovať hojenie čriev.
Odstránenie problému
Prvým krokom v programe 4Rs je odstránenie škodlivých patogénov a spúšťačov zápalov, ktoré sú spojené so zvýšenou priepustnosťou čriev. Spúšťače ako stres a chronická konzumácia alkoholu môžu telu jednotlivca narobiť veľa škody. Takže zameranie sa na tieto škodlivé faktory z tela je liečiť ho liekmi, antibiotikami, doplnkami a odporúča sa odstrániť zápalové potraviny zo stravy, vrátane:
- Alkohol
- Lepok
– Potravinové prísady
– Škroby
- Určité mastné kyseliny
– Niektoré potraviny, na ktoré je človek citlivý
Výmena živín
Druhým krokom programu 4R je nahradenie živín, ktoré spôsobujú črevné problémy, prostredníctvom zápalu. Niektoré živiny môžu pomôcť znížiť zápal v čreve a zároveň zabezpečiť podporu tráviaceho traktu. Existuje niekoľko protizápalových potravín, ktoré sú výživné. Tie obsahujú:
– Potraviny s vysokým obsahom vlákniny
- Omega-3
- Olivový olej
– Huby
- Protizápalové bylinky
Existujú určité doplnky, ktoré možno použiť na podporu funkcie trávenia tým, že pomáhajú a absorbujú živiny na podporu zdravých čriev. Čo robia tráviace enzýmy, je, že pomáhajú rozkladať tuky, bielkoviny a sacharidy v čreve. To pomôže ľuďom, ktorí majú narušený tráviaci trakt, potravinovú intoleranciu alebo celiakiu. Doplnky ako doplnky žlčových kyselín môžu pomôcť pri absorpcii živín zlúčením lipidov dohromady. Štúdie uviedli že žlčové kyseliny sa používali na liečbu pečene, žlčníka a žlčovodu pri prevencii tvorby žlčových kameňov po bariatrických operáciách.
Reinokulované črevo
Tretím krokom je program 4rs na reokuláciu črevných mikróbov prospešnými baktériami na podporu zdravej funkcie čriev. Boli preukázané štúdie že probiotické doplnky sa používali na zlepšenie čreva obnovením prospešných baktérií. S týmito doplnkami poskytujú črevám zlepšenie vylučovaním protizápalových látok do tela, pomáhajú podporovať imunitný systém, menia mikrobiálne zloženie tela a znižujú priepustnosť čriev v črevnom systéme.
Od sa nachádzajú probiotiká vo fermentovaných potravinách a považujú sa za prechodné, pretože nie sú perzistentné v gastrointestinálnom trakte a sú prospešné. Prekvapivo stále majú vplyv na ľudské zdravie vďaka ovplyvňovaniu čriev produkciou vitamínov a antimikrobiálnych zlúčenín, čím zabezpečujú rozmanitosť a funkciu čriev.
Oprava čreva
Posledným krokom programu 4Rs je oprava čreva. Tento krok zahŕňa opravu črevnej výstelky čreva pomocou špecifických živín a bylín. Tieto bylinky a doplnky môžu pomôcť znížiť priepustnosť čriev a zápal v tele. Niektoré z týchto bylín a doplnkov zahŕňajú:
– Aloe vera
– Chioská mastichová guma
– DGL (deglycyrizinované sladké drievko)
– koreň bahniatka
- L-glutamín
- Omega-3
� Polyfenoly
- vitamín D
- Zinok
záver
Pretože mnohé faktory môžu nepriaznivo ovplyvniť tráviaci systém a môžu sa podieľať na viacerých zdravotných problémoch. Hlavným cieľom programu 4Rs je minimalizovať tieto faktory, ktoré poškodzujú črevá a znižovať zápaly a zvyšovať priepustnosť čriev. Keď sa pacientovi zoznámia s prospešnými faktormi, ktoré poskytujú 4R, môže to viesť k zdravému, zahojenému črevu. Niektorí Produkty sú tu, aby pomohli podporiť gastrointestinálny systém podporou čriev, zlepšením metabolizmu cukrov a zacielením na aminokyseliny, ktoré sú určené na podporu čriev.
Rozsah našich informácií je obmedzený na chiropraktické, muskuloskeletálne a nervové zdravotné problémy alebo články, témy a diskusie o funkčnej medicíne. Funkčné zdravotné protokoly využívame na liečbu úrazov alebo porúch pohybového aparátu. Naša kancelária sa primerane pokúsila poskytnúť podporné citácie a identifikovala relevantnú výskumnú štúdiu alebo štúdie podporujúce naše príspevky. Kópie podporných výskumných štúdií tiež sprístupňujeme predstavenstvu a/alebo verejnosti na požiadanie. Ak chcete ďalej prediskutovať vyššie uvedenú tému, neváhajte sa opýtať Dr. Alexa Jimeneza alebo nás kontaktujte na adrese 915-850-0900.
Referencie:
De Santis, Stefania a kol. �Nutričné kľúče pre moduláciu črevnej bariéry.� Hranice v imunológii, Frontiers Media SA, 7. decembra 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4670985/.
Ianiro, Gianluca a kol. �Suplementácia tráviacich enzýmov pri ochoreniach gastrointestinálneho traktu.� Súčasný metabolizmus liečiv, Bentham Science Publishers, 2016, www.ncbi.nlm.nih.gov/pmc/articles/PMC4923703/.
Mu, Qinghui a kol. �Netesné črevo ako signál nebezpečenstva pre autoimunitné ochorenia.� Hranice, Frontiers, 5. máj 2017, www.frontiersin.org/articles/10.3389/fimmu.2017.00598/full.
Rezac, Shannon a kol. �Fermentované potraviny ako dietetický zdroj živých organizmov.� Hranice v mikrobiológii, Frontiers Media SA, 24. augusta 2018, www.ncbi.nlm.nih.gov/pmc/articles/PMC6117398/.
Sander, Guy R. a kol. �Rýchle narušenie funkcie črevnej bariéry gliadínom zahŕňa zmenenú expresiu apikálnych junkčných proteínov.� FEBS Press, John Wiley & Sons, Ltd, 8. augusta 2005, febs.onlinelibrary.wiley.com/doi/full/10.1016/j.febslet.2005.07.066.
Sartor, R. Balfour. �Terapeutická manipulácia s enterickou mikroflórou pri zápalových črevných ochoreniach: antibiotiká, probiotiká a prebiotiká.� Gastroenterológia, Národná lekárska knižnica USA, máj 2004, www.ncbi.nlm.nih.gov/pubmed/15168372.
Chronická bolesť je bežným zdravotným problémom, ktorý postihuje mnoho ľudí v Spojených štátoch. Zatiaľ čo niekoľko zdravotných stavov, ako je fibromyalgia a syndróm myofasciálnej bolesti, môže spôsobiť chronickú bolesť, môže sa tiež vyvinúť v dôsledku rôznych iných zdravotných problémov. Výskumné štúdie zistili, že rozšírený zápal je hlavnou príčinou chronickej bolesti. Zápal je prirodzený obranný mechanizmus proti zraneniu, chorobe alebo infekcii. Ak však zápalový proces trvá príliš dlho, môže sa stať problematickým.
Zápal signalizuje imunitnému systému, aby hojil a opravoval poškodené tkanivo, ako aj aby sa chránil pred baktériami a vírusmi. Ako je však uvedené vyššie, chronický zápal môže spôsobiť rôzne zdravotné problémy, vrátane symptómov chronickej bolesti. Úpravy zdravého životného štýlu môžu pomôcť zvládnuť chronickú bolesť, ale najprv pochopme bežné príčiny chronickej bolesti.
Čo je akútny zápal?
Akútny zápal sa napríklad vyskytuje po zranení alebo niečom tak jednoduchom, ako je bolesť hrdla. Ide o prirodzenú reakciu s nepriaznivými účinkami, čo znamená, že pôsobí lokálne v regióne, kde sa zdravotný problém vyskytuje. Bežné príznaky akútneho zápalu zahŕňajú opuch, začervenanie, teplo, bolesť a stratu funkcie, ako uvádza National Library of Medicine. Keď sa rozvinie akútny zápal, krvné cievy sa rozšíria, čo spôsobí zvýšenie prietoku krvi a biele krvinky v poranenej oblasti podporujú zotavenie.
Počas ťažkého zápalu poškodené tkanivo uvoľňuje zlúčeniny nazývané cytokíny. Cytokíny pôsobia ako „núdzové signály“, ktoré privádzajú do ľudského tela vlastné imunitné bunky, ako aj hormóny a mnohé živiny na nápravu zdravotného problému. Okrem toho látky podobné hormónom, známe ako prostaglandíny, spôsobujú krvné zrazeniny na hojenie poškodeného tkaniva a tie môžu tiež vyvolať horúčku a bolesť ako súčasť zápalového procesu. Keď sa poškodenie alebo zranenie zotaví, zápal ustúpi.
Čo je to chronický zápal?
Na rozdiel od akútneho zápalu má chronický zápal dlhodobé účinky. Chronický zápal, tiež známy ako perzistentný zápal, spôsobuje nízke úrovne zápalu v ľudskom tele, čo dokazuje zvýšenie markerov imunitného systému umiestnených v krvi a bunkových tkanivách. Chronický zápal môže tiež spôsobiť progresiu rôznych chorôb a stavov. Zvýšené úrovne zápalu sa môžu niekedy spustiť, aj keď nedošlo k zraneniu, chorobe alebo infekcii, čo môže tiež spôsobiť reakciu imunitného systému.
V dôsledku toho by imunitný systém ľudského tela mohol začať napádať zdravé bunky, tkanivá alebo orgány. Vedci sa stále snažia pochopiť dôsledky chronického zápalu v ľudskom tele a mechanizmy podieľajúce sa na tomto prirodzenom obrannom procese. Napríklad chronický zápal je spojený s rôznymi zdravotnými problémami, ako sú srdcové choroby a mŕtvica.
Jedna teória naznačuje, že keď zápal zostáva v krvných cievach, môže to podporiť hromadenie plaku. Podľa American Heart Association alebo AHA, ak imunitný systém identifikuje plak ako cudzieho votrelca, biele krvinky sa môžu pokúsiť odlepiť plak, ktorý sa nachádza v krvi prúdiacej cez tepny. To môže vytvoriť krvnú zrazeninu, ktorá môže blokovať prietok krvi do srdca alebo mozgu, čo spôsobí, že sa stane nestabilným a praskne. Rakovina je ďalší zdravotný problém spojený s chronickým zápalom. Ďalej podľa National Cancer Institute môže byť poškodenie DNA spôsobené aj chronickým zápalom.
Pretrvávajúci zápal nízkeho stupňa často nemá žiadne príznaky, ale zdravotnícki pracovníci môžu skontrolovať prítomnosť C-reaktívneho proteínu alebo CRP, známeho ako kyselina lipoová, čo je marker zápalu nájdený v krvi. Zvýšené hladiny CRP sú spojené so zvýšeným rizikom kardiovaskulárnych ochorení. Zvýšené hladiny CRP možno nájsť pri chronických poruchách, ako je lupus alebo reumatoidná artritída.
V prípade iných chronických stavov, ako je fibromyalgia, nervový systém nadmerne reaguje na špecifickú stimuláciu, ale je to zápal, ktorý spôsobuje symptómy chronickej bolesti. Subjektívne je takmer nemožné rozlíšiť medzi chronickou bolesťou spôsobenou precitliveným nervovým systémom a chronickou bolesťou spôsobenou rozsiahlym zápalom. Okrem hľadania záchytných bodov v krvnom obehu môže chronický zápal podporovať aj výživa človeka, jeho životný štýl a vystavenie sa životnému prostrediu.
Zápal je prirodzený obranný mechanizmus imunitného systému proti zraneniu, chorobe alebo infekcii. Zatiaľ čo táto zápalová reakcia môže pomôcť pri hojení a oprave tkanív, chronický, rozšírený zápal môže spôsobiť rôzne zdravotné problémy, vrátane symptómov chronickej bolesti. Vyvážený výživa, vrátane rôznych diét a pôstu, môže pomôcť znížiť zápal. Pôst, tiež známy ako kalorické obmedzenie, podporuje bunkovú apoptózu a obnovu mitochondrií. Diéta napodobňujúca pôst, ktorá je súčasťou diétneho plánu pre dlhovekosť, je diétny program, ktorý „uvádza“ ľudské telo do stavu pôstu, aby zažil výhody tradičného pôstu. Pred dodržiavaním niektorej z diét opísaných v tomto článku sa určite poraďte s lekárom.
Dr. Alex Jimenez DC, CCST Insight
Výživa, diéty, pôst a chronická bolesť
Protizápalové diéty pozostávajú najmä z konzumácie čerstvého ovocia a zeleniny, rýb a tukov. Stredomorský diétny plán je napríklad protizápalová diéta, ktorá podporuje konzumáciu mierneho množstva orechov, konzumáciu veľmi malého množstva mäsa a pitie vína. Protizápalové časti potravín, ako sú omega-3 mastné kyseliny, chránia ľudské telo pred dakúzelník spôsobené zápalom.
Protizápalová diéta zahŕňa aj vyhýbanie sa potravinám, ktoré by mohli podporovať zápal. Ideálne je znížiť množstvo potravín, ktoré jete s vysokým obsahom trans a nasýtených tukov, ako je mäso. Okrem toho protizápalová diéta obmedzuje spotrebu rafinovaných uhľohydrátov a potravín, ako je chlieb a ryža. Tieto tiež podporujú zníženie spotreby margarínu a olejov, ktoré sú nabité omega-6 mastnými kyselinami, ako je slnečnica, svetlica a kukuričné oleje.
Pôst alebo kalorické obmedzenie je už dlho známe, že znižuje oxidačný stres a spomaľuje mechanizmy starnutia v rôznych organizmoch. Účinky hladovania zahŕňajú programovanú bunkovú smrť alebo apoptózu, transkripciu, mobilnú energetickú účinnosť, mitochondriálnu biogenézu, antioxidačné mechanizmy a cirkadiánny rytmus. Pôst tiež prispieva k mitochondriálnej autofágii, známej ako mitofágia, kde sú gény v mitochondriách stimulované k apoptóze, ktorá podporuje obnovu mitochondrií.
Prerušovaný pôst vám môže pomôcť bojovať proti zápalu, zlepšiť trávenie a predĺžiť vašu dlhovekosť. Ľudské telo je navrhnuté tak, aby bolo schopné prežiť dlhší čas bez jedla. Výskumné štúdie ukázali, že prerušovaný pôst môže mať pozitívne zmeny v celkovom zložení črevnej mikroflóry. Okrem toho môže prerušovaný pôst znížiť inzulínovú rezistenciu a zároveň zvýšiť odpoveď imunitného systému. Nakoniec, prerušovaný pôst môže podporiť produkciu látky, známej ako β-hydroxybutyrát, ktorá blokuje časť imunitného systému zapojeného do zápalových ochorení, ako aj podstatne znižuje produkciu zápalových markerov, ako sú cytokíny a C-reaktívny proteín. alebo CRP, ktoré sme už spomínali vyššie.
Longevity Diet Plan, prezentovaný v knihe Dr. Valtera Longa, eliminuje konzumáciu spracovaných potravín, ktoré môžu spôsobiť zápal, podporuje pohodu a dlhovekosť. Tento jedinečný diétny program, na rozdiel od väčšiny tradičných diét, nepodporuje chudnutie. Hoci môžete zaznamenať redukciu hmotnosti, dôraz tohto jedinečného diétneho programu je zameraný na zdravšie stravovanie. Bolo preukázané, že plán Longevity Diet Plan pomáha aktivovať obnovu založenú na kmeňových bunkách, redukovať brušný tuk a predchádzať strate kostí a svalov súvisiacim s vekom, ako aj budovať odolnosť voči rozvoju kardiovaskulárnych ochorení, Alzheimerovej choroby, cukrovky a rakoviny.
Diéta napodobňujúca pôst alebo FMD vám umožňuje zažiť výhody tradičného pôstu bez toho, aby ste telo pripravili o jedlo. Hlavným rozdielom FMD je, že namiesto úplného vylúčenia všetkého jedla na niekoľko dní alebo dokonca týždňov obmedzíte príjem kalórií iba na päť dní v mesiaci. Slintačka a krívačka sa môže praktizovať raz za mesiac, aby pomohla podporiť celkové zdravie a pohodu.
Zatiaľ čo každý môže sledovať FMD samostatne, ProLon� diéta napodobňujúca pôst ponúka 5-dňový stravovací program, ktorý bol individuálne zabalený a označený na každý deň, ktorý podáva potraviny, ktoré potrebujete pre slintačky a krívačky v presných množstvách a kombináciách. Jedálenský program sa skladá z hotových alebo ľahko pripraviteľných potravín rastlinného pôvodu, vrátane tyčiniek, polievok, snackov, doplnkov, nápojového koncentrátu a čajov. Pred spustením ProLon� diéta napodobňujúca pôst, 5-dňový stravovací program, alebo niektorú z vyššie opísaných úprav životného štýlu, nezabudnite sa porozprávať so zdravotníckym pracovníkom, aby ste zistili, ktorá liečba chronickej bolesti je pre vás vhodná.
Rozsah našich informácií je obmedzený na články, témy a diskusie z oblasti chiropraxe, zdravotných problémov chrbtice a funkčnej medicíny. Ak chcete ďalej diskutovať o vyššie uvedenej téme, neváhajte sa opýtať Dr. Alexa Jimeneza alebo nás kontaktujte na adrese 915-850-0900 .
Kurátorom je Dr. Alex Jimenez
Ďalšia téma diskusie: Akútna bolesť chrbta
Bolesti chrbta je jednou z najčastejších príčin invalidity a vymeškaných dní v práci na celom svete. Bolesti chrbta sú druhým najčastejším dôvodom návštevy lekára, prevyšujú ho iba infekcie horných dýchacích ciest. Približne 80 percent populácie aspoň raz za život zažije bolesť chrbta. Vaša chrbtica je komplexná štruktúra pozostávajúca z kostí, kĺbov, väzov a svalov, okrem iných mäkkých tkanív. Zranenia a/alebo zhoršené stavy, ako napr herné disky, môže nakoniec viesť k príznakom bolesti chrbta. Športové zranenia alebo zranenia pri automobilových nehodách sú často najčastejšou príčinou bolesti chrbta, niekedy však môžu mať aj tie najjednoduchšie pohyby bolestivé následky. Našťastie alternatívne možnosti liečby, ako je chiropraktická starostlivosť, môžu pomôcť zmierniť bolesť chrbta pomocou úprav chrbtice a manuálnych manipulácií, čo v konečnom dôsledku zlepšuje úľavu od bolesti.
XYMOGEN Exkluzívne profesionálne receptúry sú dostupné prostredníctvom vybraných licencovaných odborníkov v oblasti zdravotnej starostlivosti. Internetový predaj a zľavy na receptúry XYMOGEN sú prísne zakázané.
hrdo, Doktor Alexander Jimenez sprístupňuje receptúry XYMOGEN iba pacientom v našej starostlivosti.
Zavolajte prosím do našej kancelárie, aby sme vám pridelili lekársku konzultáciu na okamžitý prístup.
Ak ste pacientom Lekárska a chiropraktická klinika úrazov, o XYMOGEN sa môžete opýtať telefonicky 915-850-0900.
Pre vaše pohodlie a prehľad XYMOGEN produkty si prosím pozrite na nasledujúcom odkaze.*XYMOGEN-Katalóg-Stiahnuť ▼
* Všetky vyššie uvedené zásady spoločnosti XYMOGEN zostávajú prísne v platnosti.
Dodržiavanie špecifickej diéty na udržanie správnej výživy môže niekedy spôsobiť, že jedenie bude stresujúce. Prirodzené úpravy životného štýlu sú kľúčom k zmene vašich stravovacích návykov, čo vám môže pomôcť žiť dlhší a zdravší život. Plán dlhovekosti, ktorý vytvoril Dr. Valter Longo, je výberom praktických stravovacích pokynov, ktoré sa zameriavajú na zmenu vašich stravovacích návykov, aby ste dosiahli celkové zdravie a pohodu.
Pravidlá diétneho plánu pre dlhovekosť
Jednoduchým dodržiavaním nižšie uvedených výživových tipov môžete prepracovať svoj súčasný diétny plán a začať sa stravovať zdravšie bez stresu z tradičnej stravy. Plán dlhovekosti eliminuje konzumáciu spracovaných potravín, ktoré môžu spôsobiť rôzne zdravotné problémy, a zvyšuje spotrebu živín, ktoré podporujú dlhovekosť. Tento jedinečný diétny program zdieľa výsledky približne 25-ročných výskumných štúdií o jednoduchom riešení, ktoré môže ľuďom pomôcť zažiť celkovú pohodu prostredníctvom správnej výživy.
Na rozdiel od väčšiny tradičných diét však plán dlhovekosti nepodporuje chudnutie. Hoci môžete zaznamenať redukciu hmotnosti, dôraz tohto jedinečného diétneho programu je zameraný na zdravšie stravovanie. Ukázalo sa, že plán Longevity Diet Plan vám pomôže aktivovať obnovu založenú na kmeňových bunkách, schudnúť a znížiť brušný tuk, zabrániť strate kostí a svalov súvisiacim s vekom, vybudovať odolnosť voči rozvoju kardiovaskulárnych chorôb, Alzheimerovej choroby, cukrovky a rakoviny. ako predĺžiť životnosť. Nižšie zhrnieme 8 najbežnejších výživových tipov Longevity Diet Plan, ktoré vám môžu v konečnom dôsledku pomôcť predĺžiť a zdravšie žiť.
Longevity Diet Plan je jedinečný diétny program navrhnutý Dr. Valterom Longom na podporu celkového zdravia, pohody a dlhovekosti. Prostredníctvom jednoduchých úprav životného štýlu môžu ľudia zmeniť svoje stravovacie návyky a využiť mnohé zdravotné benefity tohto diétneho programu. Dodržiavaním pescatariánskej diéty a dodržiavaním Diéta napodobňujúca pôst ProLon�, okrem iných výživových tipov popísaných nižšie môžu ľudia žiť dlhšie a zdravšie. Tradičné diéty môžu byť často ťažké a stresujúce, avšak plán dlhovekosti je praktický a jedinečný diétny program, ktorý môže byť vhodný pre mnohých ľudí.
Dr. Alex Jimenez DC, CCST Insight
8 výživových tipov plánu dlhovekosti
Dodržujte Pescatarian Diet
Ako súčasť Longevity Diet Plan dodržujte pescatariánske stravovanie, ktoré je takmer 100% založené na rastlinnej a rybej báze. Tiež sa uistite, že obmedzíte konzumáciu rýb na dve alebo tri porcie každý týždeň, vyhýbajte sa rybám s vyšším obsahom ortuti, ako je tuniak, mečúň, makrela a halibut. Ak máte viac ako 65 rokov a začnete pociťovať zníženú svalovú hmotu, silu a tuk, zaraďte do svojho jedálnička viac rýb spolu s inými potravinami živočíšneho pôvodu, vrátane vajec a špecifických syrov, ako je feta alebo pecorino, a jogurtov vyrobených z kozieho mäsa. mlieko.
Nejedzte príliš veľa bielkovín
Podľa plánu Longevity Diet Plan by sme mali každý deň zjesť 0.31 až 0.36 gramu bielkovín na kilogram telesného tuku. Ak vážite 130 libier, mali by ste zjesť asi 40 až 47 gramov bielkovín deň, alebo ekvivalent 1.5 filé z lososa, 1 šálka cíceru alebo 2 1/2 šálky šošovice, z čoho 30 gramov by sa malo skonzumovať v jednom jedle. Ak vážite 200 až 220 libier, mali by ste zjesť asi 60 až 70 gramov bielkovín denne alebo ekvivalent dvoch filé z lososa, 3 1/2 šálky šošovice alebo 1 1/2 šálky cíceru. Spotreba bielkovín by sa mala zvýšiť po 65. roku života. Väčšine z nás stačí zvýšenie o 10 až 20 percent alebo o 5 až 10 gramov viac každý deň. Nakoniec, diéta dlhovekosti neobsahuje živočíšne bielkoviny, ako je červené mäso, biele mäso a hydina, s výnimkou živočíšnych bielkovín v rybách. Tento jedinečný diétny program má namiesto toho porovnateľne vysoký obsah rastlinných bielkovín, ako sú strukoviny a orechy, na optimalizáciu zdravia a pohody.
Zvýšte dobré tuky a komplexné sacharidy
Ako súčasť plánu dlhovekosti by ste mali jesť vyššie množstvá polynenasýtených tukov, ako sú tie, ktoré sa nachádzajú v lososovi, mandliach, vlašských orechoch a olivovom oleji, zatiaľ čo by ste mali jesť nižšie množstvá nasýtených, hydrogenovaných a trans-tukov. Podobne ako súčasť plánu dlhovekosti by ste mali jesť aj komplexné sacharidy, ktoré sa nachádzajú v celozrnnom chlebe, strukovinách a zelenine. Uistite sa, že obmedzíte jedenie cestovín, ryže, chleba, ovocia a ovocných štiav, ktoré sa môžu premeniť na cukry, kým sa dostanú do vášho čreva.
Užívajte doplnky stravy
Ľudské telo potrebuje na správne fungovanie bielkoviny, esenciálne mastné kyseliny ako omega-3 a omega-6, vitamíny, minerály a dokonca aj cukry. Kedykoľvek sa váš príjem určitých živín stane príliš nízkym, opravné, náhradné a obranné metódy ľudského tela sa môžu spomaliť alebo zastaviť, čo umožní hubám, baktériám a vírusom spôsobiť poškodenie, ktoré môže viesť k rôznym zdravotným problémom. Užívajte vitamínové a minerálne doplnky stravy, najmä omega-3, podľa odporúčania vášho lekára.
Jedzte rôzne jedlá zo svojho Arodový pôvod
Aby ste prijali všetky potrebné živiny, musíte jesť širokú škálu jedál, no najlepšie je vybrať si jedlá, ktoré boli bežné na stole vašich rodičov, starých rodičov a prastarých rodičov. Napríklad v mnohých krajinách severnej Európy, kde sa vo všeobecnosti konzumuje mlieko, je intolerancia laktózy pomerne zriedkavá, zatiaľ čo intolerancia laktózy je celkom bežná v krajinách južnej Európy a Ázie, kde mlieko historicky nebolo súčasťou bežnej stravy dospelých. Ak sa osoba japonského pôvodu s bydliskom v Spojených štátoch náhle rozhodne začať piť mlieko, ktoré sa pravdepodobne zriedka podávalo na jedálenskom stole ich starých rodičov, pravdepodobne mu začne byť zle. Najčastejšími problémami v týchto prípadoch sú intolerancie alebo autoimunity, ako je reakcia na potraviny bohaté na lepok, ako je chlieb a cestoviny, ktoré sa vyskytujú u ľudí s celiakiou. Hoci sú potrebné ďalšie dôkazy, je možné, že potravinová intolerancia môže súvisieť s mnohými autoimunitnými poruchami, vrátane cukrovky, kolitídy a Crohnovej choroby.
Jedzte dve jedlá denne a olovrant
Podľa plánu Longevity Diet Plan je ideálne každý deň zjesť raňajky a jedno hlavné jedlo plus výživný nízkokalorický snack s nízkym obsahom cukru. Zatiaľ čo pre niektorých ľudí môže byť odporúčané jesť tri jedlá a desiatu každý deň. Mnohé výživové smernice odporúčajú, aby sme každý deň zjedli päť až šesť jedál. Keď sa ľuďom odporúča jesť často, môže byť pre nich často ťažké regulovať príjem kalórií. Za posledných dvadsať rokov sa približne 70 percent populácie v Spojených štátoch považuje za ľudí s nadváhou alebo obezitou. Je oveľa ťažšie prejedať sa podľa plánu dlhovekosti, ak budete jesť len dve a pol jedla každý deň. Na dosiahnutie množstva, ktoré by viedlo k priberaniu na váhe, by bolo treba obrovské porcie strukovín, zeleniny a rýb. Vysoká výživnosť jedál, plus množstvo jedla, vysiela signál vášmu žalúdku a mozgu, že máte dosť jedla. Tento jeden hlavný systém jedla môže byť niekedy potrebné rozdeliť na dve jedlá, aby sa predišlo problémom s trávením. Dospelí a starší ľudia náchylní na chudnutie by mali jesť tri jedlá denne. Pre ľudí, ktorí sa snažia schudnúť, ako aj pre ľudí s nadváhou alebo obezitou by najlepšou výživovou radou bolo raňajkovať denne; dajte si večeru alebo obed, ale nie oboje, a nahraďte vynechané jedlo jedným občerstvením, ktoré obsahuje menej ako 100 kalórií a nie viac ako 3 až 5 g cukru. To, ktoré jedlo vynecháte, závisí od vášho životného štýlu, ale neodporúča sa vynechávať raňajky kvôli nepriaznivým zdravotným problémom. Výhodou vynechania obeda je viac voľného času a energie. Je tu však nevýhoda jesť veľkú večeru, najmä pre ľudí, ktorí trpia refluxom kyseliny alebo problémami so spánkom. Nevýhodou vynechávania večere je však to, že môže eliminovať spoločenské jedlo dňa.
Jedzte každý deň do 12 hodín
Ďalším bežným stravovacím návykom, ktorý si osvojili mnohí storoční ľudia, je časovo obmedzené jedenie alebo obmedzenie všetkých jedál a občerstvenia v rámci 12-hodinového okna každý deň. Účinnosť tejto metódy bola preukázaná výskumnými štúdiami na ľuďoch aj na zvieratách. Vo všeobecnosti by ste mali raňajkovať o 8:8 a potom večerať o XNUMX:XNUMX. Kratšie stravovacie okno v dĺžke desať hodín alebo menej môže byť ešte lepšie na chudnutie, ale je podstatne ťažšie ho udržať a môže zvýšiť riziko vzniku vedľajších účinkov, ako sú žlčníkové kamene, a dokonca potenciálne zvýšiť pravdepodobnosť vzniku kardiovaskulárnych ochorení. Tri až štyri hodiny pred spaním by ste nemali jesť.
Dodržujte ProLon� Fasting Mimicing Diet
Zdraví ľudia do 65 rokov by mali dodržiavať ProLon� Fasting Mimicing Diet, 5-dňový stravovací program aspoň dvakrát do roka. Slintačka a krívačka je jedným z kľúčových princípov presadzovaných Plánom dlhovekosti. Diéta napodobňujúca pôst ponúka rovnaké zdravotné výhody ako pôst bez toho, aby sa skutočne postil. Zjedením 800 až 1,100 kalórií v presných množstvách a kombináciách potravín, ktoré boli jednotlivo zabalené a označené na každý deň, môžete „uviesť“ ľudské telo do stavu pôstu. Prostredníctvom rôznych výskumných štúdií Dr. Valter Longo zistil, že zbavením tela potravy týmto spôsobom naše bunky začnú rozkladať a regenerovať naše vnútorné tkanivá prostredníctvom procesu známeho ako autofágia, zabíjanie a nahradenie alebo regenerácia poškodených buniek. Okrem toho môže pôst zvrátiť rôzne zdravotné problémy, zničiť rakovinové bunky a výrazne znížiť možnosť vzniku Alzheimerovej choroby.
S Diétnym plánom dlhovekosti prezentovaným v knihe Dr. Valtera Longa budete lepšie jesť, cítiť sa lepšie a hoci nie je navrhnutý ako plán na chudnutie, môžete dokonca zhodiť pár kíl. S týmto jedinečným diétnym programom nebudete musieť zvažovať zložité pravidlá stravovania a robiť ťažké rozhodnutia. Akonáhle sa naučíte tieto úpravy životného štýlu, budete môcť zlepšiť svoje celkové zdravie a pohodu váš dlhovekosť. rozsah našich informácií je obmedzený na chiropraktiku, zdravotné problémy chrbtice a témy funkčnej medicíny. Ak chcete ďalej diskutovať o tejto téme, neváhajte sa opýtať Dr. Alexa Jimeneza alebo nás kontaktujte na adrese 915-850-0900 .
Kurátorom je Dr. Alex Jimenez
Ďalšia téma diskusie: Akútna bolesť chrbta
Bolesti chrbta je jednou z najčastejších príčin invalidity a vymeškaných dní v práci na celom svete. Bolesti chrbta sú druhým najčastejším dôvodom návštevy lekára, prevyšujú ho iba infekcie horných dýchacích ciest. Približne 80 percent populácie aspoň raz za život zažije bolesť chrbta. Vaša chrbtica je komplexná štruktúra pozostávajúca z kostí, kĺbov, väzov a svalov, okrem iných mäkkých tkanív. Zranenia a/alebo zhoršené stavy, ako napr herné disky, môže nakoniec viesť k príznakom bolesti chrbta. Športové zranenia alebo zranenia pri automobilových nehodách sú často najčastejšou príčinou bolesti chrbta, niekedy však môžu mať aj tie najjednoduchšie pohyby bolestivé následky. Našťastie alternatívne možnosti liečby, ako je chiropraktická starostlivosť, môžu pomôcť zmierniť bolesť chrbta pomocou úprav chrbtice a manuálnych manipulácií, čo v konečnom dôsledku zlepšuje úľavu od bolesti.
XYMOGEN Exkluzívne profesionálne receptúry sú dostupné prostredníctvom vybraných licencovaných odborníkov v oblasti zdravotnej starostlivosti. Internetový predaj a zľavy na receptúry XYMOGEN sú prísne zakázané.
hrdo, Doktor Alexander Jimenez sprístupňuje receptúry XYMOGEN iba pacientom v našej starostlivosti.
Zavolajte prosím do našej kancelárie, aby sme vám pridelili lekársku konzultáciu na okamžitý prístup.
Ak ste pacientom Lekárska a chiropraktická klinika úrazov, o XYMOGEN sa môžete opýtať telefonicky 915-850-0900.
Pre vaše pohodlie a prehľad XYMOGEN produkty si prosím pozrite na nasledujúcom odkaze.*XYMOGEN-Katalóg-Stiahnuť ▼
* Všetky vyššie uvedené zásady spoločnosti XYMOGEN zostávajú prísne v platnosti.
Nástroj Find A Practitioner od IFM je najväčšou sieťou odporúčaní vo funkčnej medicíne, ktorá bola vytvorená s cieľom pomôcť pacientom nájsť praktických lekárov kdekoľvek na svete. IFM Certified Practitioners sú vo výsledkoch vyhľadávania uvedení na prvom mieste vzhľadom na ich rozsiahle vzdelanie v oblasti funkčnej medicíny