Back Clinic Podpora klinickej neurofyziológie. El Paso, TX. Hovorí chiropraktik Dr. Alexander Jimenez klinická neurofyziológia. Dr. Jimenez bude skúmať klinický význam a funkčné aktivity periférnych nervových vlákien, miechy, mozgového kmeňa a mozgu v kontexte viscerálnych a muskuloskeletálnych porúch. Pacienti získajú pokročilé znalosti o anatómii, genetike, biochémii a fyziológii bolesti vo vzťahu k rôznym klinickým syndrómom. Bude zahrnutá nutričná biochémia súvisiaca s nocicepciou a bolesťou. A dôraz sa bude klásť na implementáciu týchto informácií do terapeutických programov.
Náš tím je veľmi hrdý na to, že našim rodinám a zraneným pacientom prináša iba overené liečebné protokoly. Učením úplného holistického wellness ako životného štýlu meníme nielen životy našich pacientov, ale aj ich rodiny. Robíme to preto, aby sme mohli osloviť čo najviac obyvateľov El Paso, ktorí nás potrebujú, bez ohľadu na problémy s cenovou dostupnosťou. Pre odpovede na akékoľvek otázky, ktoré môžete mať, zavolajte Dr. Jimenezovi na číslo 915-850-0900.
"Pravidlá klinického rozhodovania, klasifikácia bolesti chrbtice a predikcia výsledku liečby: Diskusia o nedávnych správach v rehabilitačnej literatúre"
abstraktné
Pravidlá klinického rozhodovania sa čoraz častejšie vyskytujú v biomedicínskej literatúre a predstavujú jednu zo stratégií zlepšovania klinického rozhodovania s cieľom zlepšiť efektívnosť a účinnosť poskytovania zdravotnej starostlivosti. V kontexte rehabilitačného výskumu boli pravidlá klinického rozhodovania zamerané predovšetkým na klasifikáciu pacientov predpovedaním ich odpovede na liečbu na špecifické terapie. Odporúčania pre vývoj pravidiel klinického rozhodovania tradične navrhujú viackrokový proces (odvodenie, validácia, analýza dopadu) s použitím definovanej metodológie. Výskumné úsilie zamerané na vývoj pravidla klinického rozhodovania založeného na diagnóze sa odchýlilo od tejto konvencie. Nedávne publikácie v tejto línii výskumu použili modifikovanú terminologickú príručku klinického rozhodovania založenú na diagnóze. Úpravy terminológie a metodológie týkajúcej sa pravidiel klinického rozhodovania môžu lekárom sťažiť rozpoznanie úrovne dôkazov spojených s rozhodovacím pravidlom a pochopenie toho, ako by sa tieto dôkazy mali implementovať na informovanie starostlivosti o pacienta. Poskytujeme stručný prehľad vývoja pravidiel klinického rozhodovania v kontexte rehabilitačnej literatúry a dvoch špecifických článkov nedávno publikovaných v Chiropractic and Manual Therapies.
Pravidlá klinickej predikcie
Zdravotníctvo prešlo dôležitým posunom paradigmy smerom k praxi založenej na dôkazoch. Prístup, ktorý má zlepšiť klinické rozhodovanie integráciou najlepších dostupných dôkazov s klinickými odbornými znalosťami a preferenciami pacientov.
V konečnom dôsledku je cieľom praxe založenej na dôkazoch zlepšiť poskytovanie zdravotnej starostlivosti. Prevedenie vedeckých dôkazov do praxe sa však ukázalo ako náročné úsilie.
Pravidlá klinického rozhodovania (CDR), tiež známe ako pravidlá klinickej predikcie, sú v rehabilitačnej literatúre čoraz bežnejšie.
Ide o nástroje určené na informovanie pri klinickom rozhodovaní identifikáciou potenciálnych prediktorov výsledku diagnostického testu, prognózy alebo terapeutickej odpovede.
V rehabilitačnej literatúre sa CDR najčastejšie používajú na predpovedanie odpovede pacienta na liečbu. Boli navrhnuté na identifikáciu klinicky relevantných podskupín pacientov s inak heterogénnymi poruchami, ako sú nešpecifické krčné alebo nízke bolesť chrbta, čo je perspektíva, na ktorú sa chceme zamerať.
Pravidlá klinickej predikcie
Schopnosť klasifikovať alebo podskupinovať pacientov s heterogénnymi poruchami, ako je bolesť chrbtice, bola zdôraznená ako priorita výskumu a následne aj zameranie veľkého výskumného úsilia. Príťažlivosť takýchto klasifikačných prístupov spočíva v ich potenciáli na zlepšenie účinnosti a účinnosti liečby priraďovaním pacientov k optimálnym terapiám. V minulosti sa klasifikácia pacientov opierala o implicitné prístupy založené na tradícii alebo nesystematických pozorovaniach. Použitie CDR na informovanie klasifikácie je jedným z pokusov o prístup viac založený na dôkazoch, ktorý je menej závislý od nepodloženej teórie.
CDR sa vyvíjajú vo viacstupňovom procese zahŕňajúcom štúdie odvodenia, validácie a analýzy vplyvu, pričom každý z nich má definovaný účel a metodologické kritériá. Ako pri všetkých formách dôkazov používaných pri rozhodovaní o pacientoch, pozornosť venovaná vhodnej metodológii štúdie je rozhodujúca pri hodnotení potenciálnych prínosov implementácie.
Výhody pravidiel klinickej predikcie
Dokáže poňať viac faktorov, ako dokáže ľudský mozog brať do úvahy
Model CDR/CPR vždy poskytne rovnaký výsledok (matematická rovnica)
V konečnom dôsledku užitočnosť CDR nespočíva v jej presnosti, ale v jej schopnosti zlepšiť klinické výsledky a zvýšiť efektivitu starostlivosti.[15] Aj keď CDR preukáže širokú validáciu, nezaručuje to, že zmení klinické rozhodovanie alebo že zmeny, ktoré vytvorí, povedú k lepšej starostlivosti.
Zmeny, ktoré spôsobí, budú mať za následok lepšiu starostlivosť. McGinn a kol.[2] identifikovali tri vysvetlenia zlyhania CDR v tomto štádiu. Po prvé, ak je úsudok lekára taký presný ako rozhodnutie informované o CDR, jeho použitie neprináša žiadne výhody. Po druhé, aplikácia CDR môže zahŕňať ťažkopádne výpočty alebo postupy, ktoré odrádzajú lekárov od používania CDR. Po tretie, použitie CDR nemusí byť možné vo všetkých prostrediach alebo okolnostiach. Okrem toho by sme zahrnuli skutočnosť, že experimentálne štúdie môžu zahŕňať pacientov, ktorí nie sú úplne reprezentatívnymi pre tých, ktorí sa vyskytujú v bežnej starostlivosti, a že to môže obmedziť skutočnú hodnotu CDR. Preto, aby sme plne porozumeli užitočnosti CDR a jej schopnosti zlepšiť poskytovanie zdravotnej starostlivosti, je potrebné vykonať pragmatické preskúmanie jej uskutočniteľnosti a dopadu pri aplikácii v prostredí odzrkadľujúcom prax v reálnom svete. Môže sa to uskutočniť pomocou rôznych návrhov štúdií, ako sú randomizované štúdie, skupinovo randomizované štúdie alebo iné prístupy, ako je napríklad skúmanie vplyvu CDR pred a po jej implementácii.
Prevalencia klasifikačných metód u pacientov s lumbálnym postihnutím pomocou McKenzieho syndrómov, vzorcov bolesti, manipulácie a pravidiel klinickej predikcie.
Cieľom bolo (1) určiť podiel pacientov s poruchami bedrového kĺbu, ktorí by mohli byť klasifikovaní pri príjme podľa McKenzieho syndrómov (McK) a klasifikácie vzorcov bolesti (PPC) pomocou metód hodnotenia mechanickej diagnostiky a terapie (MDT), manipulácie a stabilizačnej klinickej predikcie. pravidlá (CPR) a (2) pre každú kategóriu Man CPR alebo Stab CPR určia miery prevalencie klasifikácie pomocou McK a PPC.
KPR sú sofistikované pravdepodobnostné a prognostické modely, kde je skupina identifikovaných charakteristík pacienta a klinických príznakov a symptómov štatisticky spojená so zmysluplnou predikciou výsledkov pacienta.
Výskumníci vyvinuli dve samostatné KPR na identifikáciu pacientov, ktorí by priaznivo reagovali na manipuláciu.33,34 Flynn et al. vyvinuli pôvodnú manipulačnú KPR pomocou piatich kritérií, tj žiadne symptómy pod kolenom, nedávny nástup symptómov (<16 dní), nízke skóre z dotazníka o vyhýbaní sa strachu36 pre prácu (<19), hypomobilita driekovej chrbtice a vnútorné bedrové kĺby rotácia ROM (>35 pre aspoň jeden bok).33
Flynnovu KPR následne upravili Fritz a kol. na dve kritériá, ktoré zahŕňali žiadne symptómy pod kolenom a nedávny nástup symptómov (<16 dní), ako pragmatickú alternatívu na zníženie záťaže lekára pri identifikácii pacientov v primárnej starostlivosti, ktorí s najväčšou pravdepodobnosťou reagujú na manipuláciu ťahom.34 pozitívne
Pravidlo klinickej predikcie (KPR) je kombináciou klinických nálezov, ktoré štatisticky preukázali zmysluplnú predvídateľnosť pri určovaní vybraného stavu alebo prognózy pacienta, ktorému bola poskytnutá špecifická liečba 1,2. KPR sa vytvárajú pomocou štatistických metód s viacerými premennými, sú navrhnuté tak, aby skúmali predikčnú schopnosť vybraných skupín klinických premenných3,4 a sú určené na to, aby pomohli lekárom robiť rýchle rozhodnutia, ktoré môžu byť za normálnych okolností predmetom základných skreslení5. Pravidlá sú svojou povahou algoritmické a zahŕňajú zhustené informácie, ktoré identifikujú najmenší počet štatisticky diagnostických ukazovateľov vzhľadom na cieľový stav6.
Pravidlá klinickej predikcie sa vo všeobecnosti vyvíjajú pomocou 3-krokovej metódy14. Po prvé, KPR nás odvodili prospektívne
viacrozmerných štatistických metód na skúmanie prediktívnej schopnosti vybraných skupín klinických premenných3. Druhý krok zahŕňa validáciu KPR v randomizovanej kontrolovanej štúdii, aby sa znížilo riziko, že prediktívne faktory vyvinuté počas derivačnej fázy boli vybrané náhodne14. Tretí krok zahŕňa vykonanie analýzy vplyvu s cieľom určiť, ako KPR zlepšuje starostlivosť, znižuje náklady a presne definuje cieľový cieľ14.
Hoci existuje malá diskusia o tom, že starostlivo zostavené KPR môžu zlepšiť klinickú prax, pokiaľ je mi známe, neexistujú žiadne usmernenia, ktoré by špecifikovali metodické požiadavky na KPR na infúziu do všetkých prostredí klinickej praxe. Usmernenia sú vytvorené na zlepšenie prísnosti návrhu štúdie a podávania správ. Nasledujúci úvodník načrtáva potenciálne metodologické úskalia v KPR, ktoré môžu výrazne oslabiť prenosnosť algoritmu. V oblasti rehabilitácie bola väčšina KPR predpisujúcich; preto moje komentáre odrážajú normatívne KPR.
Metodologické úskalia
KPR sú navrhnuté tak, aby špecifikovali homogénny súbor charakteristík z heterogénnej populácie prospektívne vybraných po sebe nasledujúcich pacientov5,15. Výsledná použiteľná populácia je zvyčajne malá podskupina väčšej vzorky a môže predstavovať len malé percento skutočnej dennej záťaže lekára. Usporiadanie a umiestnenie väčšej vzorky by malo byť zovšeobecniteľné15,16 a následné štúdie validity vyžadujú posúdenie KPR u rôznych skupín pacientov, v rôznych prostrediach a s typickou skupinou pacientov, ktorú vidí väčšina lekárov16. Pretože mnohé KPR sú vyvinuté na základe veľmi odlišnej skupiny, ktorá môže alebo nemusí odrážať typickú populáciu pacientov, prenosnosť spektra17 mnohých súčasných algoritmov KPR môže byť obmedzená.
Pravidlá klinickej predikcie používajú na určenie účinnosti intervencie výsledky. Opatrenia výsledku musia mať jedinú operačnú definíciu5 a vyžadujú dostatočnú schopnosť reagovať na skutočné zachytenie vhodnej zmeny stavu14; okrem toho by tieto opatrenia mali mať dobre zostavené hraničné skóre16,18 a mali by ich zbierať zaslepený administrátor15. V súčasnosti sa diskutuje o výbere vhodného skóre kotvy na meranie skutočnej zmeny19-20. Väčšina výsledkov meraní používa dotazník založený na pacientoch, ako je napríklad globálne hodnotenie zmeny skóre (GRoC), ktoré je vhodné, ak sa používa krátkodobo, ale pri dlhodobých analýzach trpí zaujatosťou pri pripomínaní si19-21.
Potenciálnou nevýhodou KPR je zlyhanie pri zachovaní kvality testov a meraní používaných ako prediktory v algoritme. Perspektívny test a opatrenia by preto mali byť počas modelovania navzájom nezávislé16; každý by sa mal vykonávať zmysluplným a prijateľným spôsobom4; lekári alebo správcovia údajov by mali byť zaslepení voči výsledkom a stavu pacienta22.
Zdroje
Potenciálne úskalia pravidiel klinickej predikcie; The Journal of Manual & Manipulative Therapy Volume 16 Číslo dva [69]
Jeffrey J Hebert a Julie M Fritz; Pravidlá klinického rozhodovania, klasifikácia bolesti chrbtice a predikcia výsledku liečby: Diskusia o nedávnych správach v rehabilitačnej literatúre
Depresia je jedným z najčastejších problémov duševného zdravia v Spojených štátoch. Súčasný výskum naznačuje, že depresia je výsledkom kombinácie genetických, biologických, ekologických a psychologických aspektov. Depresia je celosvetovo hlavnou psychiatrickou poruchou s významným ekonomickým a psychologickým tlakom na spoločnosť. Našťastie sa depresia, dokonca aj tie najťažšie prípady, dá liečiť. Čím skôr je možné začať s liečbou, tým je účinnejšia.
V dôsledku toho však existuje potreba robustných biomarkerov, ktoré pomôžu zlepšiť diagnostiku, aby sa urýchlil proces objavovania lieku a/alebo lieku pre každého pacienta s poruchou. Ide o objektívne periférne fyziologické indikátory, ktorých prítomnosť možno použiť na predpovedanie pravdepodobnosti nástupu alebo existencie depresie, stratifikáciu podľa závažnosti alebo symptomatológie, indikáciu predikcie a prognózy alebo sledovanie odpovede na terapeutické zásahy. Účelom nasledujúceho článku je ukázať najnovšie poznatky, súčasné výzvy a vyhliadky do budúcnosti týkajúce sa objavovania rôznych biomarkery na depresiu a ako môžu pomôcť zlepšiť diagnostiku a liečbu.
Biomarkery pre depresiu: Nedávne poznatky, súčasné výzvy a vyhliadky do budúcnosti
abstraktné
Množstvo výskumov zahŕňalo stovky predpokladaných biomarkerov depresie, ale ešte úplne neobjasnilo ich úlohu pri depresívnom ochorení ani nezistilo, čo je abnormálne u ktorých pacientov a ako možno biologické informácie použiť na zlepšenie diagnózy, liečby a prognózy. Tento nedostatočný pokrok je čiastočne spôsobený povahou a heterogenitou depresie v spojení s metodologickou heterogenitou v rámci výskumnej literatúry a veľkým množstvom biomarkerov s potenciálom, ktorých vyjadrenie sa často mení v závislosti od mnohých faktorov. Preskúmali sme dostupnú literatúru, ktorá naznačuje, že markery zapojené do zápalových, neurotrofických a metabolických procesov, ako aj zložky neurotransmiterov a neuroendokrinného systému, predstavujú vysoko sľubných kandidátov. Tieto môžu byť merané prostredníctvom genetických a epigenetických, transkriptomických a proteomických, metabolomických a neuroimagingových hodnotení. Teraz sa vyžaduje použitie nových prístupov a systematických výskumných programov, aby sa určilo, či a ktoré biomarkery možno použiť na predpovedanie odpovede na liečbu, stratifikáciu pacientov na špecifickú liečbu a vývoj cieľov pre nové intervencie. Dospeli sme k záveru, že existuje veľa prísľubov na zníženie bremena depresie prostredníctvom ďalšieho rozvoja a rozširovania týchto výskumných ciest.
Kľúčové slová:porucha nálady, veľká depresívna porucha, zápal, odpoveď na liečbu, stratifikácia, personalizovaná medicína
úvod
Výzvy v oblasti duševného zdravia a porúch nálady
Hoci psychiatria má bremeno súvisiace s chorobami väčšie ako ktorákoľvek iná kategória lekárskej diagnostiky1, stále je zjavný rozdiel v úcte medzi fyzickým a duševným zdravím v mnohých oblastiach vrátane financovania výskumu2 a publikácií.3 Medzi ťažkosti, ktorým duševné zdravie čelí, patrí nedostatok konsenzu o klasifikácii, diagnóze a liečbe, ktorý pramení z neúplného pochopenia procesov, ktoré sú základom týchto porúch. Toto je veľmi zrejmé pri poruchách nálady, kategórii, ktorá predstavuje najväčšiu záťaž v oblasti duševného zdravia.3 Najrozšírenejšia porucha nálady, veľká depresívna porucha (MDD), je komplexné, heterogénne ochorenie, s ktorým sa môže stretnúť až 60 % pacientov. určitý stupeň rezistencie na liečbu, ktorý predlžuje a zhoršuje epizódy.4 Pri poruchách nálady a v širšej oblasti duševného zdravia by sa výsledky liečby pravdepodobne zlepšili objavením robustných, homogénnych podtypov v rámci (a medzi nimi) diagnostických kategórií, pomocou ktorých by liečba môže byť stratifikovaná. Uvedomujúc si to, v súčasnosti prebiehajú globálne iniciatívy na vymedzenie funkčných podtypov, ako sú kritériá výskumnej oblasti.5 Tvrdilo sa, že biologické markery sú prioritnými kandidátmi na podtypovanie duševných porúch.6
Zlepšenie reakcie na liečbu depresie
Napriek širokému spektru možností liečby závažnej depresie len približne tretina pacientov s MDD dosiahne remisiu, aj keď dostávajú optimálnu antidepresívnu liečbu podľa konsenzuálnych smerníc a s použitím starostlivosti založenej na meraní, a zdá sa, že miera odpovede na liečbu klesá s každou novou liečbou .7 Okrem toho depresia rezistentná na liečbu (TRD) je spojená so zvýšeným funkčným poškodením, úmrtnosťou, morbiditou a rekurentnými alebo chronickými epizódami v dlhodobom horizonte.8,9 Dosiahnutie zlepšenia odpovede na liečbu v akomkoľvek klinickom štádiu by teda poskytlo širšie výhody pre celkové výsledky depresie. Napriek značnému zaťaženiu, ktoré možno pripísať TRD, bol výskum v tejto oblasti obmedzený. Definície TRD nie sú napriek predchádzajúcim pokusom štandardizované:4 niektoré kritériá vyžadujú iba jednu liečebnú štúdiu, ktorá nedokáže dosiahnuť 50 % zníženie skóre symptómov (z overenej miery závažnosti depresie), zatiaľ čo iné vyžadujú nedosiahnutie úplnej remisie alebo nereagovanie na aspoň dve adekvátne skúšané antidepresíva rôznych tried v rámci epizódy, ktorá sa má považovať za TRD.4,10 Okrem toho sa zlepšuje určenie štádia a predikcia rezistencie na liečbu pridaním kľúčových klinických znakov závažnosti a chronicity k počtu neúspešných liečebných postupov. .9,11 Táto nejednotnosť v definícii však robí interpretáciu výskumnej literatúry o TRD ešte zložitejšou úlohou.
Na zlepšenie odpovede na liečbu je jednoznačne užitočné identifikovať prediktívne rizikové faktory nereagovania. Boli charakterizované niektoré všeobecné prediktory TRD, vrátane nedostatočnej úplnej remisie po predchádzajúcich epizódach, komorbidnej úzkosti, suicidality a skorého nástupu depresie, ako aj osobnosti (najmä nízka extraverzia, nízka závislosť od odmeny a vysoký neurotizmus) a genetické faktory.12 Tieto zistenia sú potvrdené prehľadmi syntetizujúcimi dôkazy oddelene pre farmakologickú13 a psychologickú14 liečbu depresie. Antidepresíva a kognitívno-behaviorálne terapie vykazujú približne porovnateľnú účinnosť,15 ale vzhľadom na ich odlišné mechanizmy účinku možno očakávať, že budú mať rôzne prediktory odpovede. Zatiaľ čo trauma v ranom veku bola dlho spojená s horšími klinickými výsledkami a zníženými odpoveďami na liečbu,16 skoré indikácie naznačujú, že ľudia s traumou v detstve môžu lepšie reagovať na psychologickú ako farmakologickú liečbu.17 Napriek tomu prevláda neistota a málo personalizácie resp. stratifikácia liečby sa dostala do klinickej praxe.18
Tento prehľad sa zameriava na dôkazy podporujúce užitočnosť biomarkerov ako potenciálne užitočných klinických nástrojov na zvýšenie odpovede na liečbu depresie.
Biomarkery: Systémy a zdroje
Biomarkery poskytujú potenciálny cieľ pre identifikáciu prediktorov odpovede na rôzne intervencie.19 Doterajšie dôkazy naznačujú, že markery odrážajúce aktivitu zápalových, neurotransmiterových, neurotrofických, neuroendokrinných a metabolických systémov môžu byť schopné predpovedať výsledky duševného a fyzického zdravia u jedincov v súčasnosti s depresiou. , ale medzi zisteniami je veľa nezrovnalostí.20 V tomto prehľade sa zameriavame na týchto päť biologických systémov.
Na dosiahnutie úplného pochopenia molekulárnych dráh a ich podielu na psychiatrických poruchách sa teraz považuje za dôležité posúdiť viacero biologických úrovní, čo sa bežne nazýva „omický“ prístup.21 Obrázok 1 poskytuje zobrazenie rôznych biologické úrovne, na ktorých možno hodnotiť každý z piatich systémov, a potenciálne zdroje markerov, na ktorých je možné tieto hodnotenia vykonávať. Upozorňujeme však, že zatiaľ čo každý systém možno kontrolovať na každej úrovni omics, optimálne zdroje merania sa na každej úrovni zreteľne líšia. Napríklad neuroimaging poskytuje platformu na nepriame hodnotenie štruktúry alebo funkcie mozgu, zatiaľ čo vyšetrenie bielkovín v krvi priamo hodnotí markery. Transkriptomika22 a metabolomika23 sú čoraz populárnejšie a ponúkajú hodnotenie potenciálne obrovského počtu markerov a Human Microbiom Project sa teraz pokúša identifikovať všetky mikroorganizmy a ich genetické zloženie u ľudí.24 Nové technológie zlepšujú našu schopnosť ich merať, a to aj prostredníctvom ďalších zdrojov. ; napríklad hormóny, ako je kortizol, sa teraz môžu testovať vo vlasoch alebo nechtoch na rukách (poskytuje chronickú indikáciu) alebo v pote (poskytuje kontinuálne meranie),25 ako aj v krvi, cerebrospinálnej tekutine, moči a slinách.
Vzhľadom na množstvo predpokladaných zdrojov, úrovní a systémov zapojených do depresie nie je prekvapujúce, že rozsah biomarkerov s translačným potenciálom je rozsiahly. Najmä, keď sa zvažujú interakcie medzi markermi, je možno nepravdepodobné, že skúmanie jednotlivých biomarkerov v izolácii prinesie zistenia, ktoré sú užitočné pre zlepšenie klinickej praxe. Schmidt et al26 navrhli použitie panelov biomarkerov a následne Brand et al27 načrtli návrh panelu založený na predchádzajúcich klinických a predklinických dôkazoch pre MDD, pričom identifikujú 16 „silných“ cieľov biomarkerov, z ktorých každý je zriedkavo jediným markerom. Zahŕňajú znížený objem šedej hmoty (v hipokampe, prefrontálnom kortexe a bazálnych gangliách), zmeny cirkadiánneho cyklu, hyperkortizolizmus a iné prejavy hyperaktivácie osi hypotalamus-hypofýza-nadobličky (HPA), dysfunkciu štítnej žľazy, znížený obsah dopamínu, noradrenalínu alebo kyseliny 5-hydroxyindoloctovej zvýšený glutamát, zvýšená superoxiddismutáza a peroxidácia lipidov, oslabený cyklický adenozín 3p,5p-monofosfát a mitogénom aktivovaná proteínkinázová aktivita, zvýšené prozápalové cytokíny, zmeny tryptofánu, kynurenínu, inzulínu a špecifické genetické polymorfizmy. Tieto markery neboli dohodnuté na základe konsenzu a mohli by sa merať rôznymi spôsobmi; je jasné, že cielená a systematická práca musí riešiť túto obrovskú úlohu, aby sa preukázal ich klinický prínos.
Ciele tejto recenzie
Ako zámerne široký prehľad sa tento článok snaží určiť celkové potreby výskumu biomarkerov pri depresii a rozsah, v akom biomarkery majú skutočný translačný potenciál na zlepšenie odpovede na liečbu. Začneme diskusiou o najdôležitejších a vzrušujúcich zisteniach v tejto oblasti a nasmerujeme čitateľa na konkrétnejšie recenzie týkajúce sa relevantných markerov a porovnaní. Načrtávame súčasné výzvy, ktorým čelíme vo svetle dôkazov, v kombinácii s potrebami zníženia záťaže depresie. Nakoniec sa pozrieme dopredu na dôležité cesty výskumu na riešenie súčasných výziev a ich dôsledky pre klinickú prax.
Nedávne štatistiky
Hľadanie klinicky užitočných biomarkerov pre ľudí s depresiou prinieslo za posledné polstoročie rozsiahly výskum. Najčastejšie používané spôsoby liečby boli koncipované z monoamínovej teórie depresie; následne neuroendokrinné hypotézy získali veľkú pozornosť. V posledných rokoch najplodnejší výskum obklopoval zápalovú hypotézu depresie. Veľký počet relevantných prehľadových článkov sa však zameral na všetkých päť systémov; pozri tabuľku 1 a nižšie, kde nájdete zbierku najnovších poznatkov o systémoch biomarkerov. Zatiaľ čo sa merali na mnohých úrovniach, proteíny pochádzajúce z krvi boli preskúmané najširšie a poskytujú zdroj biomarkerov, ktorý je pohodlný, nákladovo efektívny a môže byť bližšie k translačnému potenciálu ako iné zdroje; teda podrobnejšie sú uvedené biomarkery cirkulujúce v krvi.
V nedávnom systematickom prehľade Jani et al20 skúmali biomarkery periférnej krvi na depresiu v spojení s výsledkami liečby. Z iba 14 zahrnutých štúdií (vyhľadávaných do začiatku roku 2013) sa študovalo 36 biomarkerov, z ktorých 12 bolo významnými prediktormi indexov duševnej alebo fyzickej odozvy aspoň v jednom výskume. Medzi tie, ktoré boli identifikované ako potenciálne rizikové faktory neodpovedania, patrili zápalové proteíny: nízky interleukín (IL)-12p70, pomer počtu lymfocytov a monocytov; neuroendokrinné markery (dexametazón nesupresia kortizolu, vysoký cirkulujúci kortizol, znížený hormón stimulujúci štítnu žľazu); markery neurotransmiterov (nízky serotonín a noradrenalín); metabolické (cholesterol lipoproteínov s nízkou hustotou) a neurotrofické faktory (znížený proteín B viažuci vápnik S100). Okrem toho ďalšie prehľady informovali o súvislostiach medzi ďalšími biomarkermi a výsledkami liečby.19,28�30 Stručný opis predpokladaných markerov v každom systéme je uvedený v nasledujúcich častiach a v tabuľke 2.
Zápalové nálezy pri depresii
Od Smithovho kľúčového článku, ktorý načrtáva hypotézu makrofágov,31 táto zavedená literatúra zistila zvýšené hladiny rôznych prozápalových markerov u depresívnych pacientov, ktoré boli široko preskúmané.32�37 V metaanalýzach porovnávajúcich depresívnych a zdravých bolo hodnotených dvanásť zápalových proteínov. kontrolné populácie.38�43
IL-6 (P<0.001 vo všetkých metaanalýzach; zahŕňalo 31 štúdií) a CRP (P<0.001; 20 štúdií) sa pri depresii objavujú často a spoľahlivo zvýšené.40 V skorých štúdiách bol identifikovaný zvýšený tumor nekrotizujúci faktor alfa (TNF?). (P<0.001),38 ale vzhľadom na novšie výskumy (31 štúdií) je to kvôli značnej heterogenite nepresvedčivé.40 IL-1? je ešte nepresvedčivejšie spojená s depresiou, pričom metaanalýzy naznačujú vyššie hladiny depresie (P=0.03),41 vysoké hladiny iba v európskych štúdiách42 alebo žiadne rozdiely od kontrol.40 Napriek tomu nedávny článok naznačil konkrétne translačné dôsledky pre IL- 1p,44 podporovaný mimoriadne významným účinkom zvýšeného IL-1a. ribonukleová kyselina predpovedá slabú odpoveď na antidepresíva;45 ďalšie zistenia vyššie sa týkajú cirkulujúcich cytokínov odvodených z krvi. Chemokínový monocytový chemoatraktant proteín-1 ukázal zvýšenie u depresívnych účastníkov v jednej metaanalýze.39 Interleukíny IL-2, IL-4, IL-8, IL-10 a interferón gama sa významne nelíšili medzi depresívnymi pacientmi a kontrolami v metaanalytickej úrovni, ale napriek tomu preukázali potenciál, pokiaľ ide o zmenu pri liečbe: IL-8 bol hlásený ako zvýšený u pacientov s ťažkou depresiou prospektívne a prierezovo,46 rôznych vzorcov zmien v IL-10 a interferóne gama počas liečby sa vyskytli medzi včasne reagujúcimi verzus nereagujúcimi47, zatiaľ čo IL-4 a IL-2 sa znížili v súlade s remisiou symptómov.48 V metaanalýzach sa popri liečbe preukázal malý pokles pre IL-6, IL-1?, IL- 10 a CRP.43,49,50 Okrem toho TNF? sa môže redukovať iba liečbou u respondentov a zložený markerový index môže naznačovať zvýšený zápal u pacientov, ktorí následne nereagujú na liečbu.43 Je však pozoruhodné, že takmer všetky výskumy skúmajúce zápalové proteíny a odpoveď na liečbu využívajú štúdie farmakologickej liečby . Teda aspoň niektoré zápalové zmeny počas liečby možno pravdepodobne pripísať antidepresívam. Presné zápalové účinky rôznych antidepresív ešte neboli stanovené, ale dôkazy využívajúce hladiny CRP naznačujú, že jednotlivci reagujú odlišne na špecifickú liečbu na základe základného zápalu: Harley et al51 uviedli zvýšené CRP pred liečbou, čo predpovedá slabú odpoveď na psychologickú terapiu (kognitívno-behaviorálnu alebo interpersonálnu psychoterapia), ale dobrá odpoveď na nortriptylín alebo fluoxetín; Uher et al52 replikovali toto zistenie pre nortriptylín a identifikovali opačný účinok pre escitalopram. Na rozdiel od toho, Chang et al53 zistili vyššie CRP u pacientov s včasnou odpoveďou na fluoxetín alebo venlafaxín ako u pacientov, ktorí nereagovali. Okrem toho pacienti s TRD a vysokým CRP lepšie reagovali na TNF? antagonista infliximab než tie s hladinami v normálnom rozmedzí.54
Dôkazy spolu naznačujú, že aj pri kontrole faktorov, ako je index telesnej hmotnosti (BMI) a vek, sa zápalové reakcie zdajú aberantné približne u jednej tretiny pacientov s depresiou.55,56 Zápalový systém je však mimoriadne zložitý a existuje množstvo biomarkerov reprezentujúcich rôzne aspekty tohto systému. Nedávno priniesli ďalšie nové cytokíny a chemokíny dôkaz o abnormalitách depresie. Patria sem: makrofágový inhibičný proteín 1a, IL-1a, IL-7, IL-12p70, IL-13, IL-15, eotaxín, faktor stimulujúci kolónie granulocytov a makrofágov,57 IL-5,58 IL-16,59 IL- 17,60 monocytový chemoatraktant proteín-4,61 týmus a aktiváciou regulovaný chemokín,62 eotaxín-3, TNFb,63 proteín indukovaný interferónom gama 10,64 sérový amyloid A,65 rozpustná intracelulárna adhézna molekula66 a rozpustná molekula adhézie vaskulárnych buniek 1.67
Zistenia rastového faktora pri depresii
Vo svetle potenciálnej dôležitosti neneurotrofných rastových faktorov (ako sú tie, ktoré súvisia s angiogenézou), odkazujeme na neurogénne biomarkery v rámci širšej definície rastových faktorov.
Neurotrofický faktor odvodený od mozgu (BDNF) je z nich najčastejšie študovaný. Viaceré metaanalýzy preukazujú zoslabenie proteínu BDNF v sére, ktoré sa podľa všetkého zvyšuje popri liečbe antidepresívami.68�71 Najnovšia z týchto analýz naznačuje, že tieto aberácie BDNF sú výraznejšie u pacientov s najťažšou depresiou, ale zdá sa, že antidepresíva zvýšiť hladiny tohto proteínu aj pri absencii klinickej remisie.70 proBDNF bol menej študovaný ako zrelá forma BDNF, ale zdá sa, že tieto dve formy sa funkčne líšia (v zmysle ich účinkov na receptory tyrozínového receptora kinázy B) a nedávne dôkazy naznačujú, že zatiaľ čo zrelý BDNF môže byť znížený pri depresii, proBDNF môže byť nadprodukovaný.72 Periférne hodnotený nervový rastový faktor bol tiež hlásený ako nižší pri depresii ako u kontrol v metaanalýze, ale nemusí byť zmenený antidepresívnou liečbou napriek tomu, že je najviac oslabený u pacientov s ťažšou depresiou.73 Podobné zistenia boli zaznamenané v metaanalýze gliových buniekneurotrofický faktor odvodený od línie.74
Vaskulárny endoteliálny rastový faktor (VEGF) má úlohu pri podpore angiogenézy a neurogenézy spolu s ďalšími členmi rodiny VEGF (napr. VEGF-C, VEGF-D) a je prísľubom pre depresiu.75 Napriek nejednotným dôkazom dve metaanalýzy majú nedávno indikovali zvýšenia VEGF v krvi depresívnych pacientov v porovnaní s kontrolami (v 16 štúdiách; P<0.001).76,77 Nízke VEGF sa však zistilo v TRD78 a vyššie hladiny predpovedali nereakciu na liečbu antidepresívami.79 Nie je známe prečo by boli hladiny proteínu VEGF zvýšené, ale môže to byť čiastočne spôsobené prozápalovou aktivitou a/alebo zvýšením permeability hematoencefalickej bariéry v depresívnych stavoch, čo spôsobuje zníženú expresiu v mozgovomiechovom moku.80 Vzťah medzi VEGF a odpoveďou na liečbu je nejasný ; nedávna štúdia nezistila žiadny vzťah medzi sérovým VEGF alebo BDNF so závažnosťou odpovede alebo depresie, a to aj napriek poklesu popri liečbe antidepresívami.81 Inzulínu podobný rastový faktor-1 je ďalším faktorom s neurogénnymi funkciami, ktoré sa môžu zvýšiť pri depresii, čo odráža nerovnováhu v neurotrofické procesy.82,83 Základný fibroblastový rastový faktor (alebo FGF-2) je členom rodiny fibroblastových rastových faktorov a zdá sa, že je vyšší v depresívnych ako kontrolných skupinách.84 Správy však nie sú konzistentné; jeden zistil, že tento proteín bol v MDD nižší ako u zdravých kontrol, ale popri liečbe antidepresívami sa ďalej znížil.85
Ďalšie rastové faktory, ktoré neboli dostatočne preskúmané pri depresii, zahŕňajú tyrozínkinázu 2 a rozpustnú fms-podobnú tyrozínkinázu-1 (tiež nazývanú sVEGFR-1), ktoré pôsobia synergicky s VEGF, a receptory tyrozínkinázy (ktoré viažu BDNF) môžu byť oslabené pri depresii.86 Placentárny rastový faktor je tiež súčasťou rodiny VEGF, ale pokiaľ je nám známe, nebol študovaný na vzorkách so systematickou depresiou.
Zistenia metabolických biomarkerov pri depresii
Medzi hlavné biomarkery spojené s metabolickým ochorením patria leptín, adiponektín, ghrelín, triglyceridy, lipoproteín s vysokou hustotou (HDL), glukóza, inzulín a albumín.87 Vzťahy medzi mnohými z nich a depresiou boli preskúmané: leptín88 a ghrelín89 sa javia nižšie pri depresii než kontroly na periférii a môže sa zvýšiť spolu s antidepresívnou liečbou alebo remisiou. Inzulínová rezistencia môže byť zvýšená pri depresii, aj keď v malých množstvách.90 Lipidové profily, vrátane HDL-cholesterolu, sa zdajú byť zmenené u mnohých pacientov s depresiou, vrátane pacientov bez komorbidného fyzického ochorenia, hoci tento vzťah je zložitý a vyžaduje si ďalšie objasnenie.91 Okrem toho, hyperglykémia92 a hypoalbuminémia93 pri depresii boli hlásené v prehľadoch.
Vyšetrenia celkových metabolických stavov sa stávajú čoraz častejšie pomocou metabolomických panelov malých molekúl s nádejou nájsť robustný biochemický podpis pre psychiatrické poruchy. V nedávnej štúdii využívajúcej modelovanie umelej inteligencie bol súbor metabolitov ilustrujúcich zvýšenú glukózolipidovú signalizáciu vysoko prediktívny pre diagnózu MDD,94 čo podporuje predchádzajúce štúdie.95
Nálezy neurotransmiterov pri depresii
Zatiaľ čo pozornosť venovaná monoamínom pri depresii priniesla relatívne úspešnú liečbu, neboli identifikované žiadne robustné markery neurotransmiterov, ktoré by optimalizovali liečbu založenú na selektivite monoamínových cieľov antidepresív. Nedávna práca poukazuje na serotonínový (5-hydroxytryptamín) 1A receptor ako potenciálne dôležitý pre diagnózu aj prognózu depresie, kým sa nevytvoria nové genetické a zobrazovacie techniky.96 Existujú nové potenciálne liečby zamerané na 5-hydroxytryptamín; napríklad pomocou podávania 5-hydroxytryptofánu s pomalým uvoľňovaním.97 Zvýšený prenos dopamínu interaguje s inými neurotransmitermi na zlepšenie kognitívnych výsledkov, ako je rozhodovanie a motivácia.98 Podobne neurotransmitery glutamát, noradrenalín, histamín a serotonín môžu interagovať a aktivovať ako súčasť stresovej reakcie súvisiacej s depresiou; to môže znížiť produkciu 5-hydroxytryptamínu prostredníctvom „zaplavenia“. Nedávny prehľad uvádza túto teóriu a naznačuje, že pri TRD by sa to dalo zvrátiť (a obnoviť 5-HT) prostredníctvom multimodálnej liečby zameranej na viaceré neurotransmitery.99 Je zaujímavé, že k zvýšeniu serotonínu nie vždy dochádza súčasne s terapeutickými prínosmi antidepresív.100 Napriek tomu Často sa zistilo, že metabolity neurotransmiterov, ako je 3-metoxy-4-hydroxyfenylglykol, noradrenalínu alebo kyselina homovanilová, dopamínu, sa zvyšujú spolu so znížením depresie pri liečbe antidepresívami101,102, 102,103 alebo že nízke hladiny týchto metabolitov predpovedajú lepšiu odpoveď na liečba SSRI.XNUMX
Neuroendokrinné nálezy pri depresii
Kortizol je najbežnejším biomarkerom osi HPA, ktorý bol študovaný pri depresii. Početné prehľady sa zamerali na rôzne hodnotenia aktivity HPA; celkovo to naznačuje, že depresia je spojená s hyperkortizolémiou a že reakcia prebúdzania kortizolu je často oslabená.104,105 Toto je podporené nedávnym prehľadom chronických hladín kortizolu meraných vo vlasoch, čo podporuje hypotézu o hyperaktivite kortizolu pri depresii, ale o hypoaktivite pri iných ochoreniach, napr. ako panická porucha.106 Navyše najmä zvýšené hladiny kortizolu môžu predpovedať horšiu odpoveď na psychologickú107 a antidepresívnu108 liečbu. Historicky najsľubnejším neuroendokrinným markerom prospektívnej odpovede na liečbu bol test supresie dexametazónu, kde je nesupresia kortizolu po podaní dexametazónu spojená s nižšou pravdepodobnosťou následnej remisie. Tento jav sa však nepovažoval za dostatočne robustný na klinickú aplikáciu. Zistilo sa, že súvisiace markery hormón uvoľňujúci kortikotropín a hormón adrenokortikotropín, ako aj vazopresín, sú pri depresii nadmerne produkované a dehydroepiandrosterón je oslabený; pomer kortizolu k dehydroepiandrosterónu môže byť zvýšený ako relatívne stabilný marker v TRD, pretrvávajúci aj po remisii.109 Neuroendokrinné hormonálne dysfunkcie sú dlho spojené s depresiou a hypotyreóza môže tiež zohrávať kauzálnu úlohu pri depresívnej nálade.110 Okrem toho môžu reakcie štítnej žľazy normalizovať úspešnou liečbou depresie.111
V rámci vyššie uvedeného je dôležité zvážiť aj signálne dráhy naprieč systémami, ako je glykogénsyntáza kináza-3, mitogénom aktivovaná proteínkináza a cyklický adenozín 3a,5a-monofosfát, ktoré sa podieľajú na synaptickej plasticite112 a modifikujú antidepresíva.113 Ďalej potenciálni kandidáti biomarkerov, ktorí pokrývajú najmä biologické systémy, sa merajú pomocou neurozobrazovania alebo genetiky. V reakcii na nedostatok robustných a zmysluplných genómových rozdielov medzi depresívnou a nedepresívnou populáciou114 by sa mohli ukázať ako užitočnejšie nové genetické prístupy, ako sú polygénne skóre115 alebo dĺžka telomér116,117. Ďalšie biomarkery, ktoré získavajú na popularite, skúmajú cirkadiánne cykly alebo chronobiologické biomarkery využívajúce rôzne zdroje. Aktigrafia môže poskytnúť objektívne hodnotenie aktivity spánku a bdenia a odpočinku prostredníctvom akcelerometra a aktigrafické zariadenia môžu stále viac merať ďalšie faktory, ako je vystavenie svetlu. To môže byť užitočnejšie na detekciu ako bežne používané subjektívne správy o pacientoch a mohlo by to poskytnúť nové prediktory odpovede na liečbu.118 Otázka, ktoré biomarkery sú najsľubnejšie na translačné použitie, je náročná a rozvedená nižšie.
Súčasné výzvy
Pre každý z týchto piatich skúmaných neurobiologických systémov sa dôkazy riadia podobným príbehom: existuje veľa biomarkerov, ktoré sú v niektorých ohľadoch spojené s depresiou. Tieto markery sú často vzájomne prepojené komplexným a ťažko modelovateľným spôsobom. Dôkazy sú nekonzistentné a je pravdepodobné, že niektoré sú epifenomény iných faktorov a niektoré sú dôležité len pre podskupinu pacientov. Biomarkery sú pravdepodobne užitočné rôznymi spôsobmi (napr. tie, ktoré predpovedajú následnú odpoveď na liečbu, tie, ktoré naznačujú, že špecifické liečby budú pravdepodobnejšie účinné, alebo tie, ktoré sa menia s intervenciami bez ohľadu na klinické zlepšenia). Na maximalizáciu konzistentnosti a klinickej použiteľnosti biologických hodnotení v psychiatrickej populácii sú potrebné nové metódy.
Variabilita biomarkerov
Variácie biomarkerov v priebehu času a naprieč situáciami sa viac týkajú niektorých typov (napr. proteomika) ako iných (genomika). Štandardizované normy pre mnohé neexistujú alebo neboli široko akceptované. Vplyv environmentálnych faktorov na markery skutočne často závisí od genetického zloženia a iných fyziologických rozdielov medzi ľuďmi, ktoré nemožno všetky vysvetliť. To sťažuje interpretáciu hodnotenia aktivity biomarkerov a identifikácie biologických abnormalít. Kvôli množstvu potenciálnych biomarkerov sa mnohé nemerali široko alebo v úplnom paneli spolu s inými relevantnými markermi.
Bolo hlásených mnoho faktorov, ktoré menia hladiny proteínov v biologických systémoch u pacientov s afektívnymi poruchami. Spolu s faktormi súvisiacimi s výskumom, ako je trvanie a podmienky skladovania (ktoré môžu spôsobiť degradáciu niektorých zlúčenín), medzi ne patrí meraný čas dňa, etnická príslušnosť, cvičenie,119 strava (napr. aktivita mikrobiómov, najmä za predpokladu, že väčšina štúdií krvných biomarkerov to robí nevyžadujú vzorku nalačno),120 fajčenie a užívanie návykových látok,121 ako aj zdravotné faktory (ako sú sprievodné zápalové, srdcovocievne alebo iné telesné ochorenia). Napríklad, hoci sa zvýšený zápal pozoruje u depresívnych, ale inak zdravých jedincov v porovnaní so skupinami bez depresie, depresívni jedinci, ktorí majú aj komorbidný stav súvisiaci s imunitou, majú často ešte vyššie hladiny cytokínov ako tí, ktorí nemajú depresiu alebo chorobu.122 Niektoré významné faktory s pravdepodobné zapojenie do vzťahu medzi biomarkermi, depresiou a odpoveďou na liečbu je uvedené nižšie.
Stres. Endokrinné aj imunitné reakcie majú dobre známu úlohu pri reakcii na stres (fyziologický alebo psychologický) a prechodný stres v čase odberu biologických vzoriek sa vo výskumných štúdiách meria zriedkavo napriek variabilite tohto faktora medzi jednotlivcami, ktorá môže byť zvýraznená súčasnými depresívne symptómy. Akútne aj chronické psychologické stresory pôsobia ako imunitná výzva, ktorá zvýrazňuje zápalové reakcie v krátkodobom a dlhodobom horizonte.123,124 Toto zistenie sa rozširuje na skúsenosť so stresom v ranom veku, ktorý je spojený so zápalovými eleváciami u dospelých, ktoré sú nezávislé od stresu prežívaného ako dospelý.125,126 Počas traumatického zážitku v detstve bol zvýšený zápal tiež hlásený len u tých detí, ktoré boli v súčasnosti depresívne.127 Naopak, ľudia s depresiou a anamnézou traumy z detstva môžu mať otupené reakcie kortizolu na stres v porovnaní s tými, ktorí mali depresiu a bez traumy v ranom veku.128 Stresom vyvolané zmeny osi HPA sa zdajú byť vo vzájomnej súvislosti s kognitívnou funkciou,129 ako aj s podtypom depresie alebo variáciou v génoch súvisiacich s HPA.130 Stres má tiež krátkodobé a dlhodobé zhoršujúce účinky na neurogenézu131 a iné nervové 132 Nie je presne jasné, ako trauma z detstva ovplyvňuje biologické markery u dospelých s depresiou s, ale je možné, že stres v ranom veku predisponuje niektorých jedincov k pretrvávajúcim stresovým reakciám v dospelosti, ktoré sú zosilnené psychologicky a/alebo biologicky.
Kognitívne fungovanie. Neurokognitívne dysfunkcie sa často vyskytujú u ľudí s afektívnymi poruchami, dokonca aj u neliečených MDD.133 Kognitívne deficity sa zdajú byť kumulatívne popri rezistencii na liečbu.134 Neurobiologicky hrá kľúčovú úlohu v tomto vzťahu pravdepodobne os HPA129 a neurotrofické systémy135. Neurotransmitery noradrenalín a dopamín sú pravdepodobne dôležité pre kognitívne procesy, ako je učenie a pamäť.136 Zvýšené zápalové reakcie sú spojené s kognitívnym poklesom a pravdepodobne ovplyvňujú kognitívne funkcie v depresívnych epizódach,137 av remisii prostredníctvom rôznych mechanizmov.138 Skutočne, Krogh et al139 navrhli, že CRP užšie súvisí s kognitívnou výkonnosťou než s hlavnými symptómami depresie.
Vek, pohlavie a BMI. Absencia alebo prítomnosť a smer biologických rozdielov medzi mužmi a ženami boli v doterajších dôkazoch obzvlášť variabilné. Neuroendokrinné hormonálne variácie medzi mužmi a ženami interagujú s náchylnosťou na depresiu.140 Prehľad štúdií zápalu uvádza, že kontrola na základe veku a pohlavia neovplyvnila rozdiely v kontrole medzi pacientmi v zápalových cytokínoch (hoci súvislosť medzi IL-6 a depresiou sa znižovala so zvyšujúcim sa vekom, čo je v súlade s teóriami, že zápal sa vo všeobecnosti zvyšuje s vekom).41,141 77 Rozdiely VEGF medzi pacientmi a kontrolami sú väčšie v štúdiách hodnotiacich mladšie vzorky, zatiaľ čo pohlavie, BMI a klinické faktory neovplyvnili tieto porovnania na metaanalytickej úrovni.41 Zdá sa, že nedostatočná úprava BMI pri predchádzajúcich vyšetreniach zápalu a depresie mätie vysoko významné rozdiely hlásené medzi týmito skupinami.142 Definitívne sa preukázalo, že zväčšené tukové tkanivo stimuluje produkciu cytokínov a je tiež úzko spojené s metabolickými markermi.XNUMX Pretože psychotropné lieky môžu byť spojené s wei zvýšenie telesnej hmotnosti a vyššie BMI, a tieto boli spojené s rezistenciou na liečbu depresie, toto je dôležitá oblasť, ktorú treba preskúmať.
Lieky. Mnohé štúdie biomarkerov depresie (prierezové aj pozdĺžne) zhromaždili základné vzorky u neliečených účastníkov, aby sa znížila heterogenita. Mnohé z týchto hodnotení sa však robia po období vymývania z liečby, čo zanecháva potenciálne významný mätúci faktor reziduálnych zmien vo fyziológii, umocnený širokou škálou dostupných liečebných postupov, ktoré mohli mať rôzne účinky na zápal. Niektoré štúdie vylúčili psychotropné, ale nie iné užívanie liekov: najmä perorálna antikoncepčná tableta je často povolená účastníkom výskumu a nie je kontrolovaná v analýzach, čo bolo nedávno indikované na zvýšenie hladín hormónov a cytokínov.143,144 Niekoľko štúdií naznačuje, že antidepresíva lieky majú účinky na zápalovú odpoveď,34,43,49,145�147 HPA-os,108 neurotransmiter,148 a neurotrofickú149 aktivitu. Avšak, početné potenciálne liečby depresie majú odlišné a komplexné farmakologické vlastnosti, čo naznačuje, že môžu existovať diskrétne biologické účinky rôznych možností liečby, podporené súčasnými údajmi. Predpokladalo sa, že okrem monoamínových účinkov, špecifické lieky zacielené na serotonín (tj SSRI) sa pravdepodobne zameriavajú na posun Th2 pri zápale a noradrenergné antidepresíva (napr. SNRI) ovplyvňujú posun Th1 Zatiaľ nie je možné určiť účinky jednotlivých alebo kombinovaných liekov na biomarkery. Tieto sú pravdepodobne sprostredkované inými faktormi vrátane dĺžky liečby (niekoľko štúdií hodnotí dlhodobé užívanie liekov), heterogenitou vzorky a nie stratifikáciou účastníkov podľa odpovede na liečbu.
Heterogenita
metodologické. Ako už bolo spomenuté vyššie, rozdiely (medzi štúdiami av rámci nich), pokiaľ ide o to, aké liečby (a kombinácie), ktoré účastníci užívajú a užívali predtým, nevyhnutne zavedú heterogenitu do zistení výskumu, najmä do výskumu biomarkerov. Okrem toho sa mnohé ďalšie charakteristiky dizajnu a vzorky v rámci štúdií líšia, čím sa zväčšujú ťažkosti s interpretáciou a pripisovaním zistení. Patria sem parametre merania biomarkerov (napr. testovacie súpravy) a metódy zberu, uchovávania, spracovania a analýzy markerov v depresii. Hiles et al141 skúmali niektoré zdroje nekonzistentnosti v literatúre o zápaloch a zistili, že presnosť diagnostiky depresie, BMI a komorbidných ochorení boli najdôležitejšie pri hodnotení periférneho zápalu medzi depresívnymi a nedepresívnymi skupinami.
Klinické. Rozsiahla heterogenita depresívnych populácií je dobre zdokumentovaná151 a je kritickým prispievateľom k protichodným zisteniam v rámci výskumnej literatúry. Je pravdepodobné, že aj v rámci diagnóz sú abnormálne biologické profily obmedzené na podskupiny jedincov, ktoré nemusia byť stabilné v priebehu času. Súdržné podskupiny ľudí trpiacich depresiou možno identifikovať prostredníctvom kombinácie psychologických a biologických faktorov. Nižšie uvádzame potenciál na skúmanie podskupín pri riešení problémov, ktoré predstavuje variabilita a heterogenita biomarkerov.
Podtypy v rámci depresie
Doteraz žiadne homogénne podskupiny v rámci epizód alebo porúch depresie neboli schopné spoľahlivo rozlíšiť medzi pacientmi na základe prejavov symptómov alebo schopnosti reagovať na liečbu.152 Existencia podskupiny, v ktorej sú biologické aberácie výraznejšie, by pomohla vysvetliť heterogenitu medzi predchádzajúcimi štúdiami a by mohla katalyzovať cestu k stratifikovanej liečbe. Kunugi et al153 navrhli súbor štyroch potenciálnych podtypov založených na úlohe rôznych neurobiologických systémov vykazujúcich klinicky relevantné podtypy pri depresii: tie s hyperkortizolizmom prejavujúcim sa melancholickou depresiou alebo hypokortizolizmom odrážajúcim atypický podtyp, podskupinu pacientov súvisiacu s dopamínom, ktorí môžu prítomná výrazne s anhedóniou (a mohla by dobre reagovať napr. na aripiprazol) a zápalovým podtypom charakterizovaným zvýšeným zápalom. Mnoho článkov zameraných na zápal špecifikovalo prípad existencie „zápalového podtypu“ v rámci depresie.55,56,154,155 Klinické koreláty zvýšeného zápalu ešte nie sú určené a bolo urobených len málo priamych pokusov zistiť, ktorí účastníci môžu zahŕňať túto kohortu. Bolo navrhnuté, že ľudia s atypickou depresiou by mohli mať vyššie úrovne zápalu ako melancholický podtyp,156 čo možno nie je v súlade so zisteniami týkajúcimi sa osi HPA u melancholických a atypických podtypov depresie. TRD37 alebo depresia s výraznými somatickými symptómami157 sa tiež považuje za potenciálny zápalový podtyp, ale neurovegetatívne (spánok, chuť do jedla, strata libida), nálada (vrátane nízkej nálady, suicidality a podráždenosti) a kognitívne symptómy (vrátane afektívnej zaujatosti a viny)158 všetky Zdá sa, že súvisia s biologickými profilmi. Ďalší potenciálni kandidáti na zápalový podtyp zahŕňajú skúsenosti so symptómami podobnými chorobnému správaniu159,160 alebo metabolickým syndrómom.158
Sklon k (hypo)mánii môže biologicky odlíšiť pacientov trpiacich depresiou. Dôkazy teraz naznačujú, že bipolárne choroby sú mnohostrannou skupinou porúch nálady, pričom bipolárna subsyndromálna porucha sa vyskytuje častejšie, ako sa predtým rozpoznalo.161 Nepresná a/alebo oneskorená detekcia bipolárnej poruchy bola nedávno zdôraznená ako hlavný problém v klinickej psychiatrii. priemerný čas na opravu diagnózy často presahuje desaťročie162 a toto oneskorenie spôsobuje väčšiu závažnosť a náklady na celkové ochorenie.163 Keďže väčšina pacientov s bipolárnou poruchou sa spočiatku prejavovala jednou alebo viacerými depresívnymi epizódami a unipolárna depresia bola najčastejšou chybnou diagnózou, identifikácia faktory, ktoré by mohli rozlišovať medzi unipolárnou a bipolárnou depresiou, majú podstatné dôsledky.164 Poruchy bipolárneho spektra pravdepodobne neboli odhalené v niektorých predchádzajúcich výskumoch MDD biomarkerov a zhluky dôkazov naznačujú diferenciáciu aktivity osi HPA109 alebo zápalu165,166 medzi bipolárnou a unipolárnou depr. esencia. Tieto porovnania sú však vzácne, majú malú veľkosť vzorky, identifikované nevýznamné trendové účinky alebo regrutované populácie, ktoré neboli dobre charakterizované diagnózou. Tieto výskumy tiež neskúmajú úlohu reakcie na liečbu v týchto vzťahoch.
Bipolárne poruchy167 aj rezistencia na liečbu168 nie sú dichotomickými konštruktmi a ležia na kontinuách, čo zvyšuje problém identifikácie podtypu. Okrem podtypovania stojí za zmienku, že mnohé biologické abnormality pozorované pri depresii sa podobne nachádzajú aj u pacientov s inými diagnózami. Potenciálne dôležité sú teda aj transdiagnostické vyšetrenia.
Výzvy merania biomarkerov
Výber biomarkerov. Veľký počet potenciálne užitočných biomarkerov predstavuje výzvu pre psychobiológiu pri určovaní toho, ktoré markery sa akým spôsobom a pre koho podieľajú. Aby sa zvýšila výzva, relatívne málo z týchto biomarkerov bolo podrobených dostatočnému skúmaniu depresie a pre väčšinu nie sú ich presné úlohy v zdravých a klinických populáciách dobre pochopené. Napriek tomu sa uskutočnilo množstvo pokusov navrhnúť sľubné panely biomarkerov. Okrem 16 sád markerov so silným potenciálom od Branda a spol.27 Lopresti a spol. načrtávajú ďalší rozsiahly súbor markerov oxidačného stresu s potenciálom na zlepšenie odpovede na liečbu.28 Papakostas a spol. definovali a priori súbor deviatich sérových markerov zahŕňajúcich biologické systémy (BDNF, kortizol, rozpustný TNFa receptor typu II, alfa1 antitrypsín, apolipoproteín CIII, epidermálny rastový faktor, myeloperoxidáza, prolaktín a rezistín) pri validácii a replikačných vzorkách s MDD. Po spojení bolo zložené meranie týchto úrovní schopné rozlíšiť medzi MDD a kontrolnými skupinami s presnosťou 80 % – 90 %.169 Navrhujeme, aby ani tieto nepokrývali všetkých potenciálnych kandidátov v tejto oblasti; pozri tabuľku 2, kde je neúplné vymedzenie biomarkerov s potenciálom pre depresiu, ktoré obsahujú tie, ktoré majú dôkazovú základňu, ako aj sľubné nové markery.
Technology. Vďaka technologickému pokroku je teraz možné (naozaj pohodlné) merať veľké množstvo biomarkerov súčasne pri nižších nákladoch a s vyššou citlivosťou, ako tomu bolo predtým. V súčasnosti je táto schopnosť merať početné zlúčeniny pred našou schopnosťou efektívne analyzovať a interpretovať údaje, 170 niečo, čo bude pokračovať s nárastom biomarkerových polí a nových markerov, ako je metabolomika. Je to z veľkej časti spôsobené nedostatočným pochopením presných úloh a vzájomných vzťahov medzi markermi a nedostatočným pochopením toho, ako sa súvisiace markery spájajú na rôznych biologických úrovniach (napr. genetické, transkripčné, proteínové) v rámci jednotlivcov a medzi nimi. Veľké dáta využívajúce nové analytické prístupy a štandardy pomôžu pri riešení tohto problému a navrhujú sa nové metodológie; jedným príkladom je vývoj štatistického prístupu založeného na analýze založenej na toku s cieľom objaviť nové potenciálne metabolické markery založené na ich reakciách medzi sieťami a integrovať génovú expresiu s údajmi o metabolitoch.171 Techniky strojového učenia sa už používajú a pomôžu s modelmi využívajúcimi biomarkery údaje na predpovedanie výsledkov liečby v štúdiách s veľkými údajmi.172
Agregácia biomarkerov. Súčasné skúmanie radu biomarkerov je alternatívou k prehliadke izolovaných markerov, ktorá by mohla poskytnúť presnejší pohľad na komplexnú sieť biologických systémov alebo sietí.26 Tiež na pomoc pri odhaľovaní kontrastných dôkazov v doterajšej literatúre (najmä tam, kde siete biomarkerov a interakcie sú dobre pochopené), údaje biomarkerov sa potom môžu agregovať alebo indexovať. Jednou z výziev je identifikácia optimálneho spôsobu vykonávania, čo si môže vyžadovať vylepšenia technológie a/alebo nové analytické techniky (pozri časť „Veľké údaje“). Z historického hľadiska priniesli pomery medzi dvoma odlišnými biomarkermi zaujímavé zistenia.109,173 Urobilo sa len niekoľko pokusov agregovať údaje o biomarkeroch vo väčšom meradle, ako napríklad tie, ktoré využívajú analýzu hlavných komponentov sietí prozápalových cytokínov.174 V metaanalýze boli prozápalové cytokíny prevedené na skóre veľkosti s jedným účinkom pre každú štúdiu a celkovo vykazovali výrazne vyšší zápal pred liečbou antidepresívami, čo predpovedalo následnú nereakciu v ambulantných štúdiách. Kompozitné panely biomarkerov sú výzvou aj príležitosťou pre budúci výskum na identifikáciu zmysluplných a spoľahlivých zistení, ktoré možno použiť na zlepšenie výsledkov liečby.43 Štúdia Papakostasa a kol. zvolila alternatívny prístup, pričom vybrala panel heterogénnych sérových biomarkerov (zápalových, HPA os a metabolické systémy), o ktorých sa v predchádzajúcej štúdii ukázalo, že sa líšia medzi depresívnymi a kontrolnými jedincami, a zložili ich do rizikového skóre, ktoré sa líšilo v dvoch nezávislých vzorkách a kontrolnej skupine s >80 % senzitivitou a špecifickosťou.169
Veľké dáta. Použitie veľkých údajov je pravdepodobne potrebné na riešenie súčasných výziev načrtnutých okolo heterogenity, variability biomarkerov, identifikácie optimálnych markerov a privedenia tejto oblasti k translačnému aplikovanému výskumu depresie. Ako je však načrtnuté vyššie, prináša to technologické a vedecké výzvy.175 Zdravotné vedy začali využívať analýzu veľkých dát len nedávno, približne o desaťročie neskôr ako v podnikateľskom sektore. Štúdie ako iSPOT-D152 a konzorciá ako Psychiatric Genetics Consortium176 však napredujú s naším chápaním biologických mechanizmov v psychiatrii. Algoritmy strojového učenia sa vo veľmi malom počte štúdií začali používať na biomarkery depresie: nedávny výskum zhromaždil údaje od > 5,000 250 účastníkov 21 biomarkerov; po viacnásobnom imputovaní údajov sa uskutočnila regresia zosilnená strojovým učením, čo naznačuje 177 potenciálnych biomarkerov. Po ďalších regresných analýzach sa vybrali tri biomarkery, ktoré sa najsilnejšie spájajú s depresívnymi symptómami (veľmi variabilná veľkosť červených krviniek, hladiny glukózy v sére a bilirubínu). Autori dospeli k záveru, že veľké dáta možno efektívne využiť na generovanie hypotéz.XNUMX V súčasnosti prebiehajú rozsiahlejšie projekty fenotypovania biomarkerov, ktoré pomôžu posunúť našu cestu do budúcnosti neurobiológie depresie.
Vyhliadky do budúcnosti
Identifikácia panelu biomarkerov
Doterajšie zistenia v literatúre vyžadujú replikáciu vo veľkých štúdiách. To platí najmä pre nové biomarkery, ako je chemokínový týmus a aktiváciou regulovaný chemokín a rastový faktor tyrozínkináza 2, ktoré podľa našich vedomostí neboli skúmané v klinicky depresívnych a zdravých kontrolných vzorkách. Štúdie veľkých údajov musia testovať komplexné panely biomarkerov a používať sofistikované analytické techniky na úplné zistenie vzťahov medzi markermi a tými faktormi, ktoré ich modifikujú v klinických a neklinických populáciách. Okrem toho by rozsiahle replikácie analýzy hlavných komponentov mohli vytvoriť vysoko korelované skupiny biomarkerov a mohli by tiež informovať o použití „kompozitov“ v biologickej psychiatrii, čo môže zvýšiť homogenitu budúcich zistení.
Objav homogénnych podtypov
Pokiaľ ide o výber biomarkerov, môže byť potrebných viacero panelov pre rôzne potenciálne cesty, ktoré by výskum mohol zahŕňať. Súhrnne povedané, súčasné dôkazy naznačujú, že profily biomarkerov sú určite, ale nejasne zmenené v subpopulácii jedincov, ktorí v súčasnosti trpia depresiou. Môže to byť stanovené v rámci alebo medzi diagnostickými kategóriami, čo by predstavovalo určitú nekonzistentnosť zistení, ktoré možno pozorovať v tejto literatúre. Kvantifikácia biologickej podskupiny (alebo podskupín) môže byť najúčinnejšie uľahčená rozsiahlou zhlukovou analýzou panelov siete biomarkerov v depresii. To by ilustrovalo variabilitu v rámci populácie; analýzy latentných tried by mohli vykazovať odlišné klinické charakteristiky založené napríklad na zápale.
Špecifické liečebné účinky na zápal a odpoveď
Všetky bežne predpisované spôsoby liečby depresie by sa mali komplexne posúdiť z hľadiska ich špecifických biologických účinkov, pričom treba zohľadniť aj účinnosť liečebných pokusov. To môže umožniť konštruktom týkajúcim sa biomarkerov a prezentácií symptómov predpovedať výsledky rôznych antidepresív personalizovanejším spôsobom a môže to byť možné v kontexte unipolárnej aj bipolárnej depresie. To bude pravdepodobne užitočné pre nové potenciálne liečby, ako aj pre v súčasnosti indikované liečby.
Prospektívne stanovenie odpovede na liečbu
Použitie vyššie uvedených techník pravdepodobne povedie k zlepšenej schopnosti prospektívne predpovedať rezistenciu na liečbu. K tomu môžu prispieť autentickejšie a trvalejšie (napr. dlhodobé) merania odpovede na liečbu. Posúdenie iných platných meraní pohody pacienta (ako je kvalita života a každodenné fungovanie) by mohlo poskytnúť holistickejšie hodnotenie výsledku liečby, ktoré sa môže užšie spájať s biomarkermi. Zatiaľ čo samotná biologická aktivita nemusí byť schopná rozlíšiť tých, ktorí reagujú na liečbu, od tých, ktorí nereagujú, súbežné meranie biomarkerov s psychosociálnymi alebo demografickými premennými by mohlo byť integrované s informáciami o biomarkeroch pri vývoji prediktívneho modelu nedostatočnej odpovede na liečbu. Ak sa vyvinie spoľahlivý model na predpovedanie odpovede (buď pre depresívnu populáciu alebo subpopuláciu) a spätne sa validuje, translačný dizajn môže preukázať jeho použiteľnosť vo veľkej kontrolovanej štúdii.
Smerom k stratifikovanej liečbe
V súčasnosti nie sú pacienti s depresiou systematicky smerovaní k tomu, aby dostávali optimalizovaný intervenčný program. Ak sa overí, stratifikovaný dizajn štúdie by sa mohol použiť na testovanie modelu na predpovedanie nereagovania a/alebo na určenie, kde je potrebné pacienta triediť v modeli stupňovitej starostlivosti. To by mohlo byť užitočné v štandardizovaných aj naturalistických liečebných prostrediach pri rôznych typoch intervencií. Nakoniec by sa mohol vyvinúť klinicky životaschopný model, ktorý by jednotlivcom poskytol najvhodnejšiu liečbu, rozpoznal tých, u ktorých je pravdepodobné, že sa u nich vyvinie refraktérna depresia, a týmto pacientom poskytne zvýšenú starostlivosť a monitorovanie. Pacientom, u ktorých sa zistí riziko rezistencie na liečbu, môže byť predpísaná súbežná psychologická a farmakologická liečba alebo kombinovaná farmakoterapia. Ako špekulatívny príklad možno uviesť, že účastníci bez zvýšenia prozápalových cytokínov môžu byť indikovaní, aby dostávali skôr psychologickú než farmakologickú terapiu, zatiaľ čo podskupina pacientov s obzvlášť vysokým zápalom by mohla dostávať protizápalové činidlo na zvýšenie štandardnej liečby. Podobne ako pri stratifikácii môžu byť v budúcnosti možné personalizované stratégie výberu liečby. Napríklad konkrétny depresívny jedinec môže mať výrazne vysoký TNF? hladiny, ale žiadne iné biologické abnormality, a mohla by mať prospech z krátkodobej liečby TNF? Personalizovaná liečba môže zahŕňať aj monitorovanie expresie biomarkerov počas liečby s cieľom informovať o možných intervenčných zmenách, dĺžke potrebnej pokračovacej terapie alebo na zistenie skorých markerov relapsu.
Nové liečebné ciele
Existuje obrovské množstvo potenciálnych liečebných postupov, ktoré by mohli byť účinné pri depresii, ktoré neboli dostatočne preskúmané, vrátane nových alebo prehodnotených intervencií z iných medicínskych odborov. Niektoré z najpopulárnejších cieľov boli v protizápalových liekoch, ako je celekoxib (a iné inhibítory cyklooxygenázy-2), TNF? antagonisty etanercept a infliximab, minocyklín alebo aspirín. Zdá sa, že sú sľubné.178 Antiglukokortikoidné zlúčeniny vrátane ketokonazolu179 a metyrapónu180 boli skúmané na depresiu, ale obe majú nevýhody s profilom vedľajších účinkov a klinický potenciál metyrapónu je neistý. Mifepriston181 a kortikosteroidy fludrokortizón a spironolaktón182 a dexametazón a hydrokortizón183 môžu byť tiež krátkodobo účinné pri liečbe depresie. Zacielenie na antagonisty glutamátového N-metyl-d-aspartátového receptora, vrátane ketamínu, môže predstavovať účinnú liečbu depresie.184 Omega-3 polynenasýtené mastné kyseliny ovplyvňujú zápalovú a metabolickú aktivitu a zdá sa, že vykazujú určitú účinnosť pri depresii.185 Je možné, že statíny môžu majú antidepresívne účinky186 prostredníctvom relevantných neurobiologických dráh.187
Týmto spôsobom sa biochemické účinky antidepresív (pozri časť �Medikácia�) využili na klinické prínosy v iných odboroch: najmä pri gastroenterologických, neurologických a nešpecifických symptómových ochoreniach.188 Protizápalové účinky antidepresív môžu predstavovať súčasť mechanizmu tieto výhody. Lítium tiež znižuje zápal, kriticky prostredníctvom dráh glykogénsyntázy kinázy-3 Zameranie sa na tieto účinky by sa mohlo ukázať ako informatívne pre podpis biomarkerov depresie a biomarkery by zase mohli predstavovať náhradné markery pre vývoj nových liekov.
Pohľad Dr. Alexa Jimeneza
Depresia je porucha duševného zdravia charakterizovaná závažnými symptómami, ktoré ovplyvňujú náladu, vrátane straty záujmu o aktivity. Nedávne výskumné štúdie však zistili, že je možné diagnostikovať depresiu nielen pomocou symptómov správania pacienta. Podľa vedcov je identifikácia ľahko dostupných biomarkerov, ktoré by mohli presnejšie diagnostikovať depresiu, základom pre zlepšenie celkového zdravia a pohody pacienta. Napríklad klinické zistenia naznačujú, že jedinci s veľkou depresívnou poruchou alebo MDD majú nižšie hladiny molekuly acetyl-L-karnitínu alebo LAC v krvi ako zdravé kontroly. V konečnom dôsledku by stanovenie biomarkerov depresie mohlo potenciálne pomôcť lepšie určiť, kto je vystavený riziku vzniku poruchy, ako aj pomôcť zdravotníckym pracovníkom určiť najlepšiu možnosť liečby pre pacienta s depresiou.
záver
Literatúra uvádza, že približne dve tretiny pacientov s depresiou nedosiahnu remisiu počiatočnej liečby a že pravdepodobnosť neodpovede sa zvyšuje s počtom skúšaných liečebných postupov. Poskytovanie neefektívnych terapií má zásadné dôsledky na individuálne a spoločenské náklady vrátane pretrvávajúcich ťažkostí a zlého blahobytu, rizika samovrážd, straty produktivity a plytvania zdrojmi zdravotnej starostlivosti. Rozsiahla literatúra o depresii naznačuje obrovské množstvo biomarkerov s potenciálom zlepšiť liečbu ľudí s depresiou. Okrem neurotransmiterov a neuroendokrinných markerov, ktoré sú predmetom rozsiahlych štúdií po mnoho desaťročí, najnovšie poznatky zdôrazňujú zápalovú reakciu (a imunitný systém vo všeobecnosti), metabolické a rastové faktory, ktoré sú dôležité pri depresii. Nadmerné kontrastné dôkazy však ilustrujú, že existuje množstvo výziev, ktoré je potrebné vyriešiť predtým, ako sa bude môcť použiť výskum biomarkerov, aby sa zlepšil manažment a starostlivosť o ľudí s depresiou. Kvôli úplnej zložitosti biologických systémov sú súčasné vyšetrenia komplexného rozsahu markerov vo veľkých vzorkách významným prínosom pri objavovaní interakcií medzi biologickými a psychologickými stavmi u jednotlivcov. Optimalizácia merania neurobiologických parametrov a klinických meraní depresie pravdepodobne uľahčí lepšie pochopenie. Tento prehľad tiež zdôrazňuje dôležitosť skúmania potenciálne modifikujúcich faktorov (ako je choroba, vek, kognícia a lieky) pri získavaní koherentného chápania biológie depresie a mechanizmov rezistencie na liečbu. Je pravdepodobné, že niektoré markery budú najsľubnejšie na predpovedanie odpovede na liečbu alebo rezistencie na špecifickú liečbu v podskupine pacientov a súčasné meranie biologických a psychologických údajov môže zlepšiť schopnosť prospektívne identifikovať tých, ktorí sú ohrození zlými výsledkami liečby. Vytvorenie panelu biomarkerov má dôsledky na zvýšenie presnosti diagnostiky a prognózy, ako aj na individualizáciu liečby v najskoršom praktickom štádiu depresívnej choroby a na vývoj účinných nových cieľov liečby. Tieto dôsledky môžu byť obmedzené na podskupiny pacientov s depresiou. Cesty k týmto možnostiam dopĺňajú nedávne výskumné stratégie na užšie prepojenie klinických syndrómov so základnými neurobiologickými substrátmi.6 Okrem zníženia heterogenity to môže uľahčiť posun smerom k rovnosti úcty medzi fyzickým a duševným zdravím. Je jasné, že hoci je potrebné veľa práce, vytvorenie vzťahu medzi relevantnými biomarkermi a depresívnymi poruchami má podstatné dôsledky na zníženie záťaže depresie na individuálnej a spoločenskej úrovni.
Poďakovanie
Táto správa predstavuje nezávislý výskum financovaný Centrom biomedicínskeho výskumu Národného inštitútu pre výskum zdravia (NIHR) v južnom Londýne a nadáciou Maudsley NHS Foundation Trust a King's College London. Vyjadrené názory sú názormi autorov a nie nevyhnutne názormi NHS, NIHR alebo Ministerstva zdravotníctva.
poznámky pod čiarou
Zverejnenie. AHY získala za posledné 3 roky honoráre za prejav od spoločností Astra Zeneca (AZ), Lundbeck, Eli Lilly, Sunovion; honoráre za poradenstvo od Allergan, Livanova a Lundbeck, Sunovion, Janssen; a podpora výskumných grantov od Janssen a agentúr financujúcich Spojené kráľovstvo (NIHR, MRC, Wellcome Trust). AJC získala v posledných 3 rokoch honoráre za prejav od Astra Zeneca (AZ), honoráre za poradenstvo od Allergan, Livanova a Lundbeck a grantovú podporu na výskum od Lundbecku a finančných agentúr Spojeného kráľovstva (NIHR, MRC, Wellcome Trust).
Autori neuvádzajú v tejto práci žiadne iné konflikty záujmov.
Záverom možno povedať,�zatiaľ čo početné výskumné štúdie našli stovky biomarkerov depresie, len málo z nich dokázalo svoju úlohu pri depresívnom ochorení alebo ako presne by sa biologické informácie dali využiť na zlepšenie diagnózy, liečby a prognózy. Vyššie uvedený článok však skúma dostupnú literatúru o biomarkeroch zapojených počas iných procesov a porovnáva klinické nálezy s nálezmi depresie. Okrem toho nové poznatky o biomarkeroch depresie môžu pomôcť lepšie diagnostikovať depresiu, aby bolo možné pokračovať v lepšej liečbe. Informácie, na ktoré odkazuje Národné centrum pre biotechnologické informácie (NCBI).�Rozsah našich informácií je obmedzený na chiroprax, ako aj na poranenia a stavy chrbtice. Ak chcete prediskutovať túto tému, neváhajte sa opýtať Dr. Jimeneza alebo nás kontaktujte na adrese�915-850-0900 .
Kurátorom je Dr. Alex Jimenez
Ďalšie témy: Bolesť chrbta
Bolesti chrbta je jednou z najčastejších príčin invalidity a vymeškaných dní v práci na celom svete. Bolesti chrbta sa v skutočnosti pripisujú ako druhý najčastejší dôvod návštevy lekára, prevyšujú ju len infekcie horných dýchacích ciest. Približne 80 percent populácie aspoň raz za život zažije nejaký typ bolesti chrbta. Chrbtica je zložitá štruktúra pozostávajúca z kostí, kĺbov, väzov a svalov, okrem iného z mäkkých tkanív. Z tohto dôvodu môžu zranenia a/alebo zhoršené stavy, ako napr herné disky, môže nakoniec viesť k príznakom bolesti chrbta. Športové zranenia alebo zranenia pri automobilových nehodách sú často najčastejšou príčinou bolesti chrbta, niekedy však môžu mať aj tie najjednoduchšie pohyby bolestivé následky. Našťastie alternatívne možnosti liečby, ako je chiropraktická starostlivosť, môžu pomôcť zmierniť bolesť chrbta pomocou úprav chrbtice a manuálnych manipulácií, čo v konečnom dôsledku zlepšuje úľavu od bolesti.
1.�Prince M, Patel V, Saxena S, a kol. Bez duševného zdravia niet zdravia.�Lancet.�2007;370(9590): 859-877.[PubMed]
2.�Kingdon D, Wykes T. Zvýšené financovanie potrebné na výskum duševného zdravia.�BMJ.�2013;346:f402.[PubMed]
3.�Vivekanantham S, Strawbridge R, Rampuri R, Ragunathan T, Young AH. Parita publikácie pre psychiatriu.�Br J Psychiatria.�2016;209(3): 257 � 261.�[PubMed]
4.�Fava M. Diagnóza a definícia depresie rezistentnej na liečbu.�Biol Psychiatria.�2003;53(8): 649 � 659.�[PubMed]
5.�Insel T, Cuthbert B, Garvey M a kol. Kritériá výskumnej oblasti (RDoC): smerom k novému klasifikačnému rámcu pre výskum duševných porúch.�Am J Psychiatria.�2010;167(7): 748 � 751.�[PubMed]
6.�Kapur S, Phillips AG, Insel TR. Prečo biologickej psychiatrii trvalo tak dlho, kým vyvinula klinické testy a čo s tým robiť.�Mol Psychiatria.�2012;17(12): 1174 � 1179.�[PubMed]
7.�Gaynes BN, Warden D, Trivedi MH, Wisniewski SR, Fava M, Rush JA. Čo nás naučil STAR*D? Výsledky z rozsiahlej praktickej klinickej štúdie u pacientov s depresiou.�Psychiatr Serv.�2009;60(11): 1439 � 1445.�[PubMed]
8.�Fekadu A, Rane LJ, Wooderson SC, Markopoulou K, Poon L, Cleare AJ. Predikcia dlhodobejšieho výsledku depresie rezistentnej na liečbu v terciárnej starostlivosti.�Br J Psychiatria.�2012;201(5): 369-375.[PubMed]
9.�Fekadu A, Wooderson SC, Markopoulo K, Donaldson C, Papadopoulos A, Cleare AJ. Čo sa stane s pacientmi s depresiou rezistentnou na liečbu? Systematický prehľad strednodobých až dlhodobých výsledkových štúdií.�J Affect Disord.�2009;116(1�2): 4�11.�[PubMed]
10.�Trivedi M. Liečebné stratégie na zlepšenie a udržanie remisie pri veľkej depresívnej poruche.�Dialógy Clin Neurosci.�2008;10(4): 377.�[Článok bez PMC]�[PubMed]
11.�Fekadu A, Wooderson SC, Markopoulou K, Cleare AJ. The Maudsley Staging Method pre depresiu rezistentnú na liečbu: predikcia dlhodobejšieho výsledku a pretrvávanie symptómov.�Psychiatria J Clin.�2009;70(7): 952 � 957.�[PubMed]
12.�Bennabi D, Aouizerate B, El-Hage W, a kol. Rizikové faktory rezistencie na liečbu pri unipolárnej depresii: systematický prehľad.�J Affect Disord.�2015;171:137[PubMed]
13.�Serretti A, Olgiati P, Liebman MN a kol. Klinická predikcia antidepresívnej odpovede pri poruchách nálady: lineárne multivariačné verzus modely neurónovej siete.�Psychiatry Res.�2007;152(2):3.[PubMed]
14.�Driessen E, Hollon SD. Kognitívno-behaviorálna terapia pre poruchy nálady: účinnosť, moderátori a mediátori.�Psychiatr Clin North Am.�2010;33(3): 537 � 555.�[Článok bez PMC]�[PubMed]
15.�Cleare A, Pariante C, Young A, a kol. Členovia Consensus Meeting Usmernenia založené na dôkazoch na liečbu depresívnych porúch antidepresívami: revízia britskej asociácie pre psychofarmakologické smernice z roku 2008.�J Psychopharmacol.�2015;29(5): 459 � 525.�[PubMed]
16.�Tunnard C, Rane LJ, Wooderson SC, a kol. Vplyv nepriazne v detstve na samovraždu a klinický priebeh depresie rezistentnej na liečbu.�J Affect Disord.�2014;152 154:122[PubMed]
17.�Nemeroff CB, Heim CM, Thase ME a kol. Rozdielne odpovede na psychoterapiu verzus farmakoterapia u pacientov s chronickými formami veľkej depresie a traumou v detstve.�Proc Natl Acad Sci USA A.�2003;100(24): 14293 � 14296.�[Článok bez PMC]�[PubMed]
18.�Nierenberg AA. Prediktory odpovede na antidepresíva všeobecné princípy a klinické dôsledky.�Psychiatr Clin North Am.�2003;26(2): 345 � 352.�[PubMed]
19.�Daj JA. Použitie biomarkerov na predpovedanie odpovede na liečbu pri veľkej depresívnej poruche: dôkazy z minulých a súčasných štúdií.�Dialógy Clin Neurosci.�2014;16(4): 539 � 544.�[Článok bez PMC]�[PubMed]
20.�Jani BD, McLean G, Nicholl BI a kol. Hodnotenie rizika a predpovedanie výsledkov u pacientov s depresívnymi symptómami: prehľad potenciálnej úlohy biomarkerov založených na periférnej krvi.�Front Hum Neurosci.�2015;9:18[Článok bez PMC]�[PubMed]
21.�Suravajhala P, Kogelman LJ, Kadarmideen HN. Multi-omická integrácia a analýza údajov pomocou systémových genomických prístupov: metódy a aplikácie v živočíšnej výrobe, zdraví a dobrých životných podmienkach zvierat.�Genet Sel Evol.�2016;48(1): 1.�[Článok bez PMC]�[PubMed]
23.�Peng B, Li H, Peng XX. Funkčná metabolomika: od objavu biomarkerov po preprogramovanie metabolómu.�Proteínová bunka.�2015;6(9): 628 � 637.�[Článok bez PMC]�[PubMed]
24.�Aagaard K, Petrosino J, Keitel W, a kol. Stratégia projektu Human Microbiom Project pre komplexný odber vzoriek ľudského mikrobiómu a prečo je to dôležité.�FASEB J.�2013;27(3): 1012-1022.[Článok bez PMC]�[PubMed]
25.�Sonner Z, Wilder E, Heikenfeld J, a kol. Mikrofluidika ekrinných potných žliaz, vrátane rozdelenia biomarkerov, transportu a biologických dôsledkov.�Biomikrofluidiká.�2015;9(3): 031301.[Článok bez PMC]�[PubMed]
26.�Schmidt HD, Shelton RC, Duman RS. Funkčné biomarkery depresie: diagnostika, liečba a patofyziológia.�Neuropsychofarm.�2011;36(12): 2375 � 2394.�[Článok bez PMC]�[PubMed]
27.�J Brand S, Moller M, H Harvey B. Prehľad biomarkerov nálady a psychotických porúch: pitva medzi klinickými a predklinickými korelátmi.�Curr Neuropharmacol.�2015;13(3): 324-368.[Článok bez PMC]�[PubMed]
28.�Lopresti AL, Maker GL, Hood SD, Drummond PD. Prehľad periférnych biomarkerov pri veľkej depresii: potenciál biomarkerov zápalového a oxidačného stresu.�Prog Neuropsychopharmacol Biol Psychiatry.�2014;48:102[PubMed]
29.�Fu CH, Steiner H, Costafreda SG. Prediktívne nervové biomarkery klinickej odpovede pri depresii: metaanalýza funkčných a štrukturálnych neuroimagingových štúdií farmakologických a psychologických terapií.�Neurobiol Dis.�2013;52:75[PubMed]
30.�Mamdani F, Berlim M, Beaulieu M, Labbe A, Merette C, Turecki G. Biomarkery génovej expresie odpovede na liečbu citalopramom pri veľkej depresívnej poruche.�Transl Psychiatria.�2011;1(6): e13.[Článok bez PMC]�[PubMed]
31.�Smith RS. Makrofágová teória depresie.�Med hypotézy.�1991;35(4): 298 � 306.�[PubMed]
32.�Irwin MR, Miller AH. Depresívne poruchy a imunita: 20 rokov pokroku a objavov.�Brain Behav Immun.�2007;21(4): 374 � 383.�[PubMed]
33.�Maes M, Leonard B, Myint A, Kubera M, Verkerk R. Nová �5-HT� hypotéza depresie: bunkami sprostredkovaná imunitná aktivácia indukuje indolamín 2,3-dioxygenázu, čo vedie k zníženiu plazmatického tryptofánu a zvýšenej syntéze škodlivé katabolity tryptofánu (TRYCAT), ktoré oba prispievajú k nástupu depresie.�Prog Neuropsychopharmacol Biol Psychiatry.�2011;35(3): 702-721.[PubMed]
34.�Miller AH, Maletic V, Raison CL. Zápal a jeho nespokojnosť: Úloha cytokínov v patofyziológii veľkej depresie.�Biol Psychiatria.�2009;65(9): 732 � 741.�[Článok bez PMC]�[PubMed]
35.�Miller AH, Raison CL. Úloha zápalu pri depresii: od evolučného imperatívu k cieľu modernej liečby.�Nat Rev Immun.�2016;16(1): 22 � 34.�[Článok bez PMC]�[PubMed]
36.�Raison CL, Capuron L, Miller AH. Cytokíny spievajú blues: zápal a patogenéza depresie.�Trends Immun.�2006;27(1): 24 � 31.�[Článok bez PMC]�[PubMed]
37.�Raison CL, Felger JC, Miller AH. Odolnosť voči zápalu a liečbe pri veľkej depresii: Dokonalá búrka.�Psychiatr Times.�2013;30(9)
38.�Dowlati Y, Herrmann N, Swardfager W, a kol. Metaanalýza cytokínov pri veľkej depresii.�Biol Psychiatria.�2010;67(5): 446 � 457.�[PubMed]
39.�Eyre HA, Air T, Pradhan A, a kol. Metaanalýza chemokínov pri veľkej depresii.�Prog Neuropsychopharmacol Biol Psychiatry.�2016;68:1[Článok bez PMC]�[PubMed]
40.�Haapakoski R, Mathieu J, Ebmeier KP, Alenius H, Kivim�ki M. Kumulatívna metaanalýza interleukínov 6 a 1?, faktor nekrózy nádorov? a C-reaktívny proteín u pacientov s veľkou depresívnou poruchou.�Brain Behav Immun.�2015;49:206[Článok bez PMC]�[PubMed]
41.�Howren MB, Lamkin DM, Suls J. Asociácie depresie s C-reaktívnym proteínom, IL-1 a IL-6: metaanalýza.�Psychosom Med.�2009;71(2): 171 � 186.�[PubMed]
42.�Liu Y, Ho RC-M, Mak A. Interleukín (IL)-6, tumor nekrotizujúci faktor alfa (TNF-a) a rozpustné receptory interleukínu-2 (sIL-2R) sú zvýšené u pacientov s veľkou depresívnou poruchou: meta- analýza a metaregresia.�J Affect Disord.�2012;139(3): 230 � 239.�[PubMed]
43.�Strawbridge R, Arnone D, Danese A, Papadopoulos A, Herane Vives A, Cleare AJ. Zápal a klinická odpoveď na liečbu depresie: metaanalýza.�Eur Neuropsychopharmacol.�2015;25(10): 1532 � 1543.�[PubMed]
44.�Farooq RK, Asghar K, Kanwal S, Zulqernain A. Úloha zápalových cytokínov pri depresii: Zameranie na interleukín-1? (recenzia)�Biomed Rep.�2017;6(1): 15 � 20.�[Článok bez PMC]�[PubMed]
45.�Cattaneo A, Ferrari C, Uher R a kol. Absolútne merania inhibičného faktora migrácie makrofágov a interleukínu-1-? Hladiny mRNA presne predpovedajú odpoveď na liečbu u depresívnych pacientov.�Int J Neuropsychopharmacol.�2016;19(10):pyw045.�[Článok bez PMC]�[PubMed]
46.�Baune B, Smith E, Reppermund S, a kol. Zápalové biomarkery predpovedajú depresívne, ale nie úzkostné symptómy počas starnutia: perspektívna štúdia pamäti a starnutia v Sydney.�Psychoneuroendokrinol.�2012;37(9): 1521 � 1530.�[PubMed]
47.�Fornaro M, Rocchi G, Escelsior A, Contini P, Martino M. Rozdielne cytokínové trendy u depresívnych pacientov užívajúcich duloxetín môžu naznačovať rozdielne biologické pozadie.�J Affect Disord.�2013;145(3): 300 � 307.�[PubMed]
48.�Hernandez ME, Mendieta D, Martinez-Fong D, a kol. Zmeny hladín cirkulujúcich cytokínov počas 52-týždňovej liečby SSRI pre veľkú depresívnu poruchu.�Eur Neuropsychopharmacol.�2008;18(12): 917 � 924.�[PubMed]
49.�Hannestad J, DellaGioia N, Bloch M. Účinok liečby antidepresívami na sérové hladiny zápalových cytokínov: metaanalýza.�Neuropsychofarmakológia.�2011;36(12): 2452-2459.[Článok bez PMC]�[PubMed]
50.�Hiles SA, Attia J, Baker AL. Zmeny interleukínu-6, C-reaktívneho proteínu a interleukínu-10 u ľudí s depresiou po liečbe antidepresívami: metaanalýza.�Brain Behav Immun; Prezentované na: 17. výročnom stretnutí PsychoNeuroImmunology Research Society PsychoNeuroImmunology: Crossing Disciplines to Combat Disease; 2012. s. S44.
51.�Harley J, Luty S, Carter J, Mulder R, Joyce P. Zvýšený C-reaktívny proteín pri depresii: Prediktor dobrého dlhodobého výsledku s antidepresívami a zlého výsledku pri psychoterapii.�J Psychopharmacol.�2010;24(4): 625 � 626.�[PubMed]
52.�Uher R, Tansey KE, Dew T, a kol. Zápalový biomarker ako diferenciálny prediktor výsledku liečby depresie escitalopramom a nortriptylínom.�Am J Psychiatria.�2014;171(2): 1278-1286.[PubMed]
53.�Chang HH, Lee IH, Gean PW a kol. Odpoveď na liečbu a kognitívne poškodenie pri veľkej depresii: Asociácia s C-reaktívnym proteínom.�Brain Behav Immun.�2012;26(1): 90 � 95.�[PubMed]
54.�Raison CL, Rutherford RE, Woolwine BJ a kol. Randomizovaná kontrolovaná štúdia antagonistu tumor nekrotizujúceho faktora infliximabu na liečbu rezistentnej depresie: úloha základných zápalových biomarkerov.�Psychiatria JAMA.�2013;70(1): 31 � 41.�[Článok bez PMC]�[PubMed]
56.�Raison CL, Miller AH. Je depresia zápalová porucha?�Curr Psychiatry Rep.�2011;13(6): 467 � 475.�[Článok bez PMC]�[PubMed]
57.�Simon N, McNamara K, Chow C, a kol. Podrobné vyšetrenie abnormalít cytokínov pri veľkej depresívnej poruche.�Eur Neuropsychopharmacol.�2008;18(3): 230 � 233.�[Článok bez PMC]�[PubMed]
58.�Dahl J, Ormstad H, Aass HC a kol. Plazmatické hladiny rôznych cytokínov sú zvýšené počas prebiehajúcej depresie a po zotavení sú znížené na normálnu úroveň.�Psychoneuroendokrinol.�2014;45:77[PubMed]
59.�Stelzhammer V, Haenisch F, Chan MK a kol. Proteomické zmeny v sére u pacientov s prvou depresiou, ktorí predtým neužívali antidepresíva.�Int J Neuropsychopharmacol.�2014;17(10): 1599 � 1608.�[PubMed]
60.�Liu Y, HO RCM, Mak A. Úloha interleukínu (IL)-17 pri úzkosti a depresii pacientov s reumatoidnou artritídou.�Int J Rheum Dis.�2012;15(2): 183 � 187.�[PubMed]
61.�Diniz BS, Sibille E, Ding Y a kol. Biologický podpis plazmy a patológia mozgu súvisiace s pretrvávajúcim kognitívnym poškodením pri depresii v neskoršom veku.�Mol Psychiatria.�2015;20(5): 594 � 601.�[Článok bez PMC][PubMed]
62.�Janelidze S., Ventorp F., Erhardt S. a kol. Zmenené hladiny chemokínov v mozgovomiechovom moku a plazme u pokusných o samovraždu.�Psychoneuroendokrinol.�2013;38(6): 853 � 862.�[PubMed]
63.�Powell TR, Schalkwyk LC, Heffernan AL, a kol. Nádorový nekrotický faktor a jeho ciele v dráhe zápalových cytokínov sú identifikované ako predpokladané transkriptomické biomarkery pre odpoveď na escitalopram.�Eur Neuropsychopharmacol.�2013;23(9): 1105 � 1114.�[PubMed]
64.�Wong M, Dong C, Maestre-Mesa J, Licinio J. Polymorfizmy v génoch súvisiacich so zápalom sú spojené s náchylnosťou na veľkú depresiu a antidepresívnu odpoveď.�Mol Psychiatria.�2008;13(8): 800 � 812.�[Článok bez PMC]�[PubMed]
65.�Kling MA, Alesci S, Csako G, a kol. Pretrvávajúci prozápalový stav nízkeho stupňa u neliečených, remitovaných žien s veľkou depresívnou poruchou, čo dokazujú zvýšené sérové hladiny C-reaktívneho proteínu akútnej fázy a sérového amyloidu A.�Biol Psychiatria.�2007;62(4): 309 � 313.�[Článok bez PMC][PubMed]
66.�Schaefer M, Sarkar S, Schwarz M, Friebe A. Rozpustná intracelulárna adhézna molekula-1 u pacientov s unipolárnymi alebo bipolárnymi afektívnymi poruchami: výsledky z pilotnej štúdie.�Neuropsychobiol.�2016;74(1): 8-14.[PubMed]
67.�Dimopoulos N, Piperi C, Salonicioti A, a kol. Zvýšenie plazmatickej koncentrácie adhéznych molekúl pri depresii v neskoršom veku.�Int J Geriatr Psychiatry.�2006;21(10): 965 � 971.�[PubMed]
68.�Bocchio-Chiavetto L, Bagnardi V, Zanardini R, et al. Hladiny BDNF v sére a plazme pri veľkej depresii: replikačná štúdia a metaanalýzy.�World J Biol Psychiatry.�2010;11(6): 763 � 773.�[PubMed]
69.�Brunoni AR, Lopes M, Fregni F. Systematický prehľad a metaanalýza klinických štúdií o veľkej depresii a hladinách BDNF: dôsledky pre úlohu neuroplasticity pri depresii.�Int J Neuropsychopharmacol.�2008;11(8): 1169 � 1180.�[PubMed]
70.�Molendijk M, Spinhoven P, Polak M, Bus B, Penninx B, Elzinga B. Sérové koncentrácie BDNF ako periférne prejavy depresie: dôkazy zo systematického prehľadu a metaanalýz na 179 asociáciách.�Mol Psychiatria.�2014;19(7): 791 � 800.�[PubMed]
71.�Sen S, Duman R, Sanacora G. Sérový neurotrofický faktor odvodený z mozgu, depresia a antidepresíva: metaanalýzy a dôsledky.�Biol Psychiatria.�2008;64(6): 527 � 532.�[Článok bez PMC][PubMed]
72.�Zhou L, Xiong J, Lim Y a kol. Upregulácia krvného proBDNF a jeho receptorov pri veľkej depresii.�J Affect Disord.�2013;150(3): 776 � 784.�[PubMed]
73.�Chen YW, Lin PY, Tu KY, Cheng YS, Wu CK, Tseng PT. Výrazne nižšie hladiny nervového rastového faktora u pacientov s veľkou depresívnou poruchou ako u zdravých jedincov: metaanalýza a systematický prehľad.�Neuropsychiatr Dis Treat.�2014;11:925[Článok bez PMC]�[PubMed]
74.�Lin PY, Tseng PT. Znížené hladiny neurotrofického faktora odvodeného od línie gliových buniek u pacientov s depresiou: metaanalytická štúdia.�J Psychiatr Res.�2015;63:20[PubMed]
75.�Warner-Schmidt JL, Duman RS. VEGF ako potenciálny cieľ pre terapeutickú intervenciu pri depresii.�Curr Op Pharmacol.�2008;8(1): 14 � 19.�[Článok bez PMC]�[PubMed]
76.�Carvalho AF, Kühler CA, McIntyre RS a kol. Periférny vaskulárny endoteliálny rastový faktor ako nový biomarker depresie: metaanalýza.�Psychoneuroendokrinol.�2015;62:18[PubMed]
77.�Tseng PT, Cheng YS, Chen YW, Wu CK, Lin PY. Zvýšené hladiny vaskulárneho endotelového rastového faktora u pacientov s veľkou depresívnou poruchou: metaanalýza.�Eur Neuropsychopharmacol.�2015;25(10): 1622 � 1630.�[PubMed]
78.�Carvalho L, Torre J, Papadopoulos A, a kol. Nedostatok klinického terapeutického prínosu antidepresív je spojený s celkovou aktiváciou zápalového systému.�J Affect Disord.�2013;148(1): 136 � 140.�[PubMed]
79.�Clark-Raymond A, Meresh E, Hoppensteadt D, a kol. Vaskulárny endoteliálny rastový faktor: Potenciálny prediktor odpovede na liečbu pri veľkej depresii.�World J Biol Psychiatry.�2015: 1�11.�[PubMed]
80.�Isung J, Mobarrez F, Nordstr�m P, �sberg M, Jokinen J. Nízky plazmatický vaskulárny endoteliálny rastový faktor (VEGF) spojený s dokončenou samovraždou.�World J Biol Psychiatry.�2012;13(6): 468 � 473.�[PubMed]
81.�Buttensch�n HN, Foldager L, Elfving B, Poulsen PH, Uher R, Mors O. Neurotrofické faktory pri depresii v reakcii na liečbu.�J Affect Disord.�2015;183:287[PubMed]
82.�Szcz?sny E, ?lusarczyk J, G?ombik K, et al. Možný príspevok IGF-1 k depresívnej poruche.�Pharmacol Rep.�2013;65(6): 1622 � 1631.�[PubMed]
83.�Tu KY, Wu MK, Chen YW a kol. Významne vyššie hladiny periférneho inzulínu podobného rastového faktora-1 u pacientov s veľkou depresívnou poruchou alebo bipolárnou poruchou ako u zdravých kontrol: metaanalýza a prehľad podľa usmernenia PRISMA.�Med.�2016;95(4):e2411.�[Článok bez PMC]�[PubMed]
84.�Wu CK, Tseng PT, Chen YW, Tu KY, Lin PY. Výrazne vyššie hladiny periférneho fibroblastového rastového faktora-2 u pacientov s veľkou depresívnou poruchou: predbežná metaanalýza podľa pokynov MOOSE.�Med.�2016;95(33):e4563.�[Článok bez PMC]�[PubMed]
85.�He S, Zhang T, Hong B, a kol. Znížené hladiny fibroblastového rastového faktora-2 v sére u pacientov s veľkou depresívnou poruchou pred a po liečbe.�Neurosci Lett.�2014;579:168[PubMed]
86.�Dwivedi Y, Rizavi HS, Conley RR, Roberts RC, Tamminga CA, Pandey GN. Zmenená génová expresia neurotrofického faktora odvodeného z mozgu a receptorovej tyrozínkinázy B v posmrtnom mozgu samovrahov.�Arch Gen Psychiatry.�2003;60(8): 804 � 815.�[PubMed]
87.�Srikanthan K, Feyh A, Visweshwar H, Shapiro JI, Sodhi K. Systematický prehľad biomarkerov metabolického syndrómu: Panel pre včasnú detekciu, manažment a stratifikáciu rizika v populácii Západnej Virgínie.�Int J Med Sci.�2016;13(1): 25.�[Článok bez PMC]�[PubMed]
88.�Lu XY. Leptínová hypotéza depresie: potenciálne spojenie medzi poruchami nálady a obezitou?�Curr Op Pharmacol.�2007;7(6): 648 � 652.�[Článok bez PMC]�[PubMed]
89.�Wittekind DA, Kluge M. Ghrelin pri psychiatrických poruchách�Prehľad.�Psychoneuroendokrinol.�2015;52:176[PubMed]
90.�Kan C, Silva N, Golden SH a kol. Systematický prehľad a metaanalýza asociácie medzi depresiou a inzulínovou rezistenciou.�Diabetes Care.�2013;36(2): 480 � 489.�[Článok bez PMC]�[PubMed]
91.�Liu X, Li J, Zheng P a kol. Plazmatická lipidomika odhaľuje potenciálne lipidové markery veľkej depresívnej poruchy.�Anal Bioanal Chem.�2016;408(23): 6497 � 6507.�[PubMed]
92.�Lustman PJ, Anderson RJ, Freedland KE, De Groot M, Carney RM, Clouse RE. Depresia a zlá kontrola glykémie: metaanalytický prehľad literatúry.�Diabetes Care.�2000;23(7): 934 � 942.�[PubMed]
93.�Maes M. Dôkaz imunitnej odpovede pri veľkej depresii: prehľad a hypotéza.�Prog NeuroPsychopharmacol Biol Psychiatry.�1995;19(1): 11 � 38.�[PubMed]
94.�Zheng H, Zheng P, Zhao L a kol. Prediktívna diagnostika veľkej depresie pomocou metabolomiky založenej na NMR a stroja na podporu vektorov najmenších štvorcov.�Clinica Chimica Acta.�2017;464:223.[PubMed]
95.�Xia Q, Wang G, Wang H, Xie Z, Fang Y, Li Y. Štúdia metabolizmu glukózy a lipidov u pacientov s depresiou v prvej epizóde.�Psychiatria J Clin.�2009;19:241.
96.�Kaufman J, DeLorenzo C, Choudhury S, Parsey RV. 5-HT1A receptor pri veľkej depresívnej poruche.�Eur Neuropsychofarmakológia.�2016;26(3): 397 � 410.�[Článok bez PMC]�[PubMed]
97.�Jacobsen JP, Krystal AD, Krishnan KRR, Caron MG. Doplnkové pomalé uvoľňovanie 5-hydroxytryptofánu pre depresiu rezistentnú na liečbu: klinické a predklinické zdôvodnenie.�Trends Pharmacol Sci.�2016;37(11): 933 � 944.�[Článok bez PMC]�[PubMed]
98.�Salamone JD, Correa M, Yohn S, Cruz LL, San Miguel N, Alatorre L. Farmakológia rozhodovacieho správania súvisiaceho s úsilím: dopamín, depresia a individuálne rozdiely.�Správanie procesov.�2016;127:3[PubMed]
99.�Coplan JD, Gopinath S, Abdallah CG, Berry BR. Neurobiologická hypotéza depresie rezistentnej na liečbu�mechanizmy pre neúčinnosť selektívneho inhibítora spätného vychytávania serotonínu�Front Behav Neurosci.�2014;8:189[Článok bez PMC]�[PubMed]
100.�Popa D, Cerdan J, Rep�rant C, a kol. Dlhodobá štúdia odtoku 5-HT počas chronickej liečby fluoxetínom pomocou novej techniky chronickej mikrodialýzy u vysoko emocionálneho kmeňa myší.�Eur J Pharmacol.�2010;628(1): 83 � 90.�[PubMed]
101.�Atake K, Yoshimura R, Hori H a kol. Duloxetín, selektívny inhibítor spätného vychytávania noradrenalínu, zvýšil plazmatické hladiny 3-metoxy-4-hydroxyfenylglykolu, ale nie kyseliny homovanilovej u pacientov s veľkou depresívnou poruchou.�Clin Psychopharmacol Neurosci.�2014;12(1): 37 � 40.�[Článok bez PMC][PubMed]
102.�Ueda N, Yoshimura R, Shinkai K, Nakamura J. Plazmatické hladiny metabolitov katecholamínov predpovedajú odpoveď na sulpirid alebo fluvoxamín pri veľkej depresii.�Farmakopsychiatria.�2002;35(05): 175-181.[PubMed]
103.�Yamana M, Atake K, Katsuki A, Hori H, Yoshimura R. Krvné biologické markery na predpovedanie odpovede na escitalopram u pacientov s veľkou depresívnou poruchou: predbežná štúdia.�J Depress Úzkosť.�2016;5: 222.
104.�Parker KJ, Schatzberg AF, Lyons DM. Neuroendokrinné aspekty hyperkortizolizmu pri veľkej depresii.�Horm Behav.�2003;43(1): 60 � 66.�[PubMed]
105.�Stetler C, Miller GE. Depresia a aktivácia hypotalamus-hypofýza-nadobličky: kvantitatívne zhrnutie štyroch desaťročí výskumu.�Psychosom Med.�2011;73(2): 114 � 126.�[PubMed]
106.�Herane Vives A, De Angel V, Papadopoulos A a kol. Vzťah medzi kortizolom, stresom a psychiatrickým ochorením: Nové poznatky pomocou analýzy vlasov.�J Psychiatr Res.�2015;70:38[PubMed]
107.�Fischer S, Strawbridge R, Vives AH, Cleare AJ. Kortizol ako prediktor odozvy psychologickej terapie pri depresívnych poruchách: systematický prehľad a metaanalýza.�Br J Psychiatria.�2017;210(2): 105 � 109.�[PubMed]
108.�Anacker C, Zunszain PA, Carvalho LA, Pariante CM. Glukokortikoidný receptor: pivot depresie a liečby antidepresívami?�Psychoneuroendokrinológia.�2011;36(3): 415 � 425.�[Článok bez PMC][PubMed]
109.�Markopoulou K, Papadopoulos A, Juruena MF, Poon L, Pariante CM, Cleare AJ. Pomer kortizolu/DHEA pri depresii rezistentnej na liečbu.�Psychoneuroendokrinol.�2009;34(1): 19 � 26.�[PubMed]
110.�Joffe RT, Pearce EN, Hennessey JV, Ryan JJ, Stern RA. Subklinická hypotyreóza, nálada a kognícia u starších dospelých: prehľad.�Int J Geriatr Psychiatry.�2013;28(2): 111 � 118.�[Článok bez PMC][PubMed]
111.�Duval F, Mokrani MC, Erb A a kol. Chronobiologický stav osi hypotalamu�hypofýzy�štítnej žľazy a výsledok antidepresív pri ťažkej depresii.�Psychoneuroendokrinol.�2015;59:71[PubMed]
112.�Marsden W. Synaptická plasticita pri depresii: molekulárne, bunkové a funkčné koreláty.�Prog Neuropsychopharmacol Biol Psychiatry.�2013;43:168[PubMed]
113.�Duman RS, Voleti B. Signálne dráhy, ktoré sú základom patofyziológie a liečby depresie: nové mechanizmy pre rýchlo pôsobiace látky.�Trendy Neurosci.�2012;35(1): 47-56.[Článok bez PMC]�[PubMed]
114.�Ripke S, Wray NR, Lewis CM a kol. Megaanalýza celogenómových asociačných štúdií pre veľkú depresívnu poruchu.�Mol Psychiatria.�2013;18(4): 497 � 511.�[Článok bez PMC]�[PubMed]
115.�Mullins N, Power R, Fisher H a kol. Polygénne interakcie s nepriaznivými vplyvmi prostredia v etiológii veľkej depresívnej poruchy.�Psychol Med.�2016;46(04): 759 � 770.�[Článok bez PMC]�[PubMed]
116.�Lewis S. Neurologické poruchy: teloméry a depresia.�Nat Rev Neurosci.�2014;15(10): 632.[PubMed]
117.�Lindqvist D, Epel ES, Mellon SH a kol. Psychiatrické poruchy a dĺžka telomér leukocytov: základné mechanizmy spájajúce duševné choroby so starnutím buniek.�Neurosci Biobehav Rev.�2015;55:333[Článok bez PMC]�[PubMed]
118.�McCall WV. Biomarker pokojovej aktivity na predpovedanie odpovede na SSRI pri veľkej depresívnej poruche.�J Psychiatr Res.�2015;64:19[Článok bez PMC]�[PubMed]
119.�Schuch FB, Deslandes AC, Stubbs B, Gosmann NP, da Silva CTB, poslanec Almeida Fleck. Neurobiologické účinky cvičenia na veľkú depresívnu poruchu: systematický prehľad.�Neurosci Biobehav Rev.�2016;61:1[PubMed]
120.�Foster JA, Neufeld K-AM. Črevná mozgová os: ako mikrobióm ovplyvňuje úzkosť a depresiu.�Trendy Neurosci.�2013;36(5): 305 � 312.�[PubMed]
121.�Quattrocki E, Baird A, Yurgelun-Todd D. Biologické aspekty spojenia medzi fajčením a depresiou.�Harv Rev Psychiatry.�2000;8(3): 99 � 110.�[PubMed]
122.�Maes M, Kubera M, Obuchowiczwa E, Goehler L, Brzeszcz J. Viacnásobné komorbidity depresie vysvetlené dráhami (neuro)zápalového a oxidačného a nitrozatívneho stresu.�Neuro Endocrinol Lett.�2011;32(1): 7 � 24.�[PubMed]
123.�Miller G, Rohleder N, Cole SW. Chronický interpersonálny stres predpovedá aktiváciu prozápalových a protizápalových signálnych dráh o šesť mesiacov neskôr.�Psychosom Med.�2009;71(1): 57.�[Článok bez PMC][PubMed]
124.�Steptoe A, Hamer M, Chida Y. Účinky akútneho psychologického stresu na cirkulujúce zápalové faktory u ľudí: prehľad a metaanalýza.�Brain Behav Immun.�2007;21(7): 901 � 912.�[PubMed]
125.�Danese A, Moffitt TE, Harrington H a kol. Nepriaznivé skúsenosti z detstva a rizikové faktory dospelých chorôb súvisiacich s vekom: depresia, zápal a zoskupovanie markerov metabolického rizika.�Arch Pediatr Adolesc Med.�2009;163(12): 1135 � 1143.�[Článok bez PMC]�[PubMed]
126.�Danese A, Pariante CM, Caspi A, Taylor A, Poulton R. Týranie v detstve predpovedá zápal dospelých v štúdii o priebehu života.�Proc Natl Acad Sci USA A.�2007;104(4): 1319 � 1324.�[Článok bez PMC][PubMed]
127.�Danese A, Caspi A, Williams B a kol. Biologické zakotvenie stresu prostredníctvom zápalových procesov v detstve.�Mol Psychiatria.�2011;16(3): 244 � 246.�[Článok bez PMC]�[PubMed]
128.�Suzuki A, Poon L, Kumari V, Cleare AJ. Strach predpojatý v emocionálnom spracovaní tváre po traume z detstva ako ukazovateľ odolnosti a zraniteľnosti voči depresii.�Týranie detí.�2015;20(4): 240 � 250.�[PubMed]
129.�Strawbridge R, Young AH. HPA os a kognitívna dysregulácia pri poruchách nálady. In: McIntyre RS, Cha DS, redaktori.�Kognitívna porucha pri závažnej depresívnej poruche: klinický význam, biologické substráty a možnosti liečby.�Cambridge: Cambridge University Press; 2016. s. 179�193.
130.�Keller J, Gomez R., Williams G a kol. HPA os pri veľkej depresii: kortizol, klinická symptomatológia a genetická variácia predpovedajú kogníciu.�Mol Psychiatria.�2016. augusta 16;�Epub.�[Článok bez PMC]�[PubMed]
131.�Hanson ND, Owens MJ, Nemeroff CB. Depresia, antidepresíva a neurogenéza: kritické prehodnotenie.�Neuropsychofarmak.�2011;36(13): 2589 � 2602.�[Článok bez PMC]�[PubMed]
132.�Chen Y, Baram TZ. Aby sme pochopili, ako stres v ranom veku preprogramuje kognitívne a emocionálne mozgové siete.�Neuropsychofarmak.�2015;41(1): 197 � 206.�[Článok bez PMC]�[PubMed]
133.�Porter RJ, Gallagher P, Thompson JM, Young AH. Neurokognitívne poškodenie u pacientov bez liekov s veľkou depresívnou poruchou.�Br J Psychiatria.�2003;182:214[PubMed]
134.�Gallagher P, Robinson L, Gray J, Young A, Porter R. Neurokognitívna funkcia po remisii pri veľkej depresívnej poruche: potenciálny objektívny marker odpovede?�Psychiatria Aust NZJ.�2007;41(1): 54 � 61.�[PubMed]
136.�B�ckman L, Nyberg L, Lindenberger U, Li SC, Farde L. Korelatívna triáda medzi starnutím, dopamínom a poznaním: súčasný stav a vyhliadky do budúcnosti.�Neurosci Biobehav Rev.�2006;30(6): 791 � 807.�[PubMed]
137.�Allison DJ, Ditor DS. Bežná zápalová etiológia depresie a kognitívnej poruchy: terapeutický cieľ.�J Neurozápal.�2014;11:151[Článok bez PMC]�[PubMed]
138.�Rosenblat JD, Brietzke E, Mansur RB, Maruschak NA, Lee Y, McIntyre RS. Zápal ako neurobiologický substrát kognitívnej poruchy pri bipolárnej poruche: dôkazy, patofyziológia a dôsledky liečby.�J Affect Disord.�2015;188:149[PubMed]
139.�Krogh J, Benros ME, J�rgensen MB, Vesterager L, Elfving B, Nordentoft M. Súvislosť medzi symptómami depresie, kognitívnymi funkciami a zápalom pri veľkej depresii.�Brain Behav Immun.�2014;35:70[PubMed]
140.�Soares CN, Zitek B. Citlivosť reprodukčných hormónov a riziko depresie v priebehu ženského životného cyklu: kontinuita zraniteľnosti?�J Psychiatria Neurosci.�2008;33(4): 331.�[Článok bez PMC]�[PubMed]
141.�Hiles SA, Baker AL, de Malmanche T, Attia J. Metaanalýza rozdielov v IL-6 a IL-10 medzi ľuďmi s depresiou a bez nej: skúmanie príčin heterogenity.�Brain Behav Immun.�2012;26(7): 1180 � 1188.�[PubMed]
142.�Fontana L, Eagon JC, Trujillo ME, Scherer PE, Klein S. Sekrécia adipokínu viscerálneho tuku je spojená so systémovým zápalom u obéznych ľudí.�Cukrovka.�2007;56(4): 1010 � 1013.�[PubMed]
143.�Divani AA, Luo X, Datta YH, Flaherty JD, Panoskaltsis-Mortari A. Účinok perorálnych a vaginálnych hormonálnych kontraceptív na zápalové krvné biomarkery.�Mediátory zápalu.�2015;2015: 379501.[Článok bez PMC]�[PubMed]
144.�Ramsey JM, Cooper JD, Penninx BW, Bahn S. Variácie v sérových biomarkeroch s pohlavím a ženským hormonálnym stavom: dôsledky pre klinické testy.�Sci Rep.�2016;6:26947[Článok bez PMC]�[PubMed]
145.�Eyre H, Lavretsky H, Kartika J, Qassim A, Baune B. Modulačné účinky tried antidepresív na vrodený a adaptívny imunitný systém pri depresii.�Farmakopsychiatria.�2016;49(3): 85-96.[Článok bez PMC]�[PubMed]
146.�Hiles SA, Baker AL, de Malmanche T, Attia J. Interleukín-6, C-reaktívny proteín a interleukín-10 po liečbe antidepresívami u ľudí s depresiou: metaanalýza.�Psychol Med.�2012;42(10): 2015 � 2026.�[PubMed]
148.�Artigas F. Serotonínové receptory zapojené do antidepresívnych účinkov.�Pharmacol Ther.�2013;137(1): 119 � 131.�[PubMed]
149.�Lee BH, Kim YK. Úloha BDNF v patofyziológii veľkej depresie a pri liečbe antidepresívami.�Psychiatrické vyšetrenie.�2010;7(4): 231 � 235.�[Článok bez PMC]�[PubMed]
150.�Hashimoto K. Zápalové biomarkery ako diferenciálne prediktory antidepresívnej odpovede.�Int J Mol Sci.�2015;16(4): 7796 � 7801.�[Článok bez PMC]�[PubMed]
151.�Goldberg D. Heterogenita „závažnej depresie“Svetová psychiatria.�2011;10(3): 226-228.[Článok bez PMC]�[PubMed]
152.�Arnow BA, Blasey C., Williams LM a kol. Podtypy depresie pri predpovedaní odpovede na antidepresíva: správa zo štúdie iSPOT-D.�Am J Psychiatria.�2015;172(8): 743 � 750.�[PubMed]
153.�Kunugi H, Hori H, Ogawa S. Biochemické markery subtypujúce veľkú depresívnu poruchu.�Psychiatria Clin Neurosci.�2015;69(10): 597 � 608.�[PubMed]
154.�Baune B., Stuart M., Gilmour A. a kol. Vzťah medzi podtypmi depresie a kardiovaskulárnym ochorením: systematický prehľad biologických modelov.�Transl Psychiatria.�2012;2(3): e92.[Článok bez PMC]�[PubMed]
155.�Vogelzangs N, Duivis HE, Beekman AT, a kol. Asociácia depresívnych porúch, charakteristík depresie a antidepresívnej medikácie so zápalom.�Transl Psychiatria.�2012;2: E79.[Článok bez PMC]�[PubMed]
156.�Lamers F, Vogelzangs N, Merikangas K, De Jonge P, Beekman A, Penninx B. Dôkazy o rozdielnej úlohe funkcie osi HPA, zápalu a metabolického syndrómu pri melancholickej verzus atypickej depresii.�Mol Psychiatria.�2013;18(6): 692 � 699.�[PubMed]
157.�Penninx BW, Milaneschi Y, Lamers F, Vogelzangs N. Pochopenie somatických dôsledkov depresie: biologické mechanizmy a úloha profilu symptómov depresie.�BMC Med.�2013;11(1): 1.[Článok bez PMC]�[PubMed]
158.�Capuron L, Su S, Miller AH, a kol. Symptómy depresie a metabolický syndróm: je zápal základným článkom?�Biol Psychiatria.�2008;64(10): 896 � 900.�[Článok bez PMC]�[PubMed]
159.�Dantzer R, O�Connor JC, Freund GG, Johnson RW, Kelley KW. Od zápalu k chorobe a depresii: keď si imunitný systém podmaňuje mozog.�Nat Rev Neurosci.�2008;9(1): 46-56.[Článok bez PMC]�[PubMed]
160.�Maes M., Berk M., Goehler L. a kol. Depresia a chorobné správanie sú Janusove reakcie na spoločné zápalové cesty.�BMC Med.�2012;10:66[Článok bez PMC]�[PubMed]
161.�Merikangas KR, Jin R, He JP a kol. Prevalencia a korelácie poruchy bipolárneho spektra vo svetovej iniciatíve prieskumu duševného zdravia.�Arch Gen Psychiatry.�2011;68(3): 241 � 251.�[Článok bez PMC][PubMed]
162.�Hirschfeld RM, Lewis L, Vornik LA. Vnímanie a vplyv bipolárnej poruchy: ako ďaleko sme sa skutočne dostali? Výsledky národného prieskumu asociácie depresív a maniodepresív z roku 2000 u jedincov s bipolárnou poruchou.�Psychiatria J Clin.�2003;64(2): 161 � 174.�[PubMed]
163.�Young AH, MacPherson H. Detekcia bipolárnej poruchy.�Br J Psychiatria.�2011;199(1): 3-4.[PubMed]
164.�V�hringer PA, Perlis RH. Rozlišovanie medzi bipolárnou poruchou a veľkou depresívnou poruchou.�Psychiatr Clin North Am.�2016;39(1): 1 � 10.�[PubMed]
165.�Becking K, Spijker AT, Hoencamp E, Penninx BW, Schoevers RA, Boschloo L. Poruchy osi hypotalamus-hypofýza-nadobličky a imunologická aktivita rozlišujúca medzi unipolárnymi a bipolárnymi depresívnymi epizódami.�PLoS One.�2015;10(7):e0133898.�[Článok bez PMC]�[PubMed]
166.�Huang TL, Lin FC. Hladiny C-reaktívneho proteínu s vysokou citlivosťou u pacientov s veľkou depresívnou poruchou a bipolárnou mániou.�Prog NeuroPsychopharmacol Biol Psychiatry.�2007;31(2): 370 � 372.�[PubMed]
167.�Angst J, Gamma A, Endrass J. Rizikové faktory pre bipolárne a depresie spektrá.�Acta Psychiatr Scand.�2003;418:15[PubMed]
168.�Fekadu A. Wooderson S., Donaldson C. a kol. Multidimenzionálny nástroj na kvantifikáciu rezistencie na liečbu depresie: Maudsleyho metóda stagingu.�Psychiatria J Clin.�2009;70(2): 177.�[PubMed]
169.�Papakostas G, Shelton R, Kinrys G, a kol. Vyhodnotenie viacnásobného biologického diagnostického testu na sérum na veľkú depresívnu poruchu: pilotná a replikačná štúdia.�Mol Psychiatria.�2013;18(3): 332 � 339.�[PubMed]
170.�Fan J, Han F, Liu H. Výzvy analýzy veľkých dát.�Natl Sci Rev.�2014;1(2): 293-314.[Článok bez PMC]�[PubMed]
171.�Li L, Jiang H, Qiu Y, Ching WK, Vassiliadis VS. Objav biomarkerov metabolitov: analýza toku a prístup siete reakcie a reakcie.�BMC Syst Biol.�2013;7(Suppl 2):S13.�[Článok bez PMC][PubMed]
172.�Patel MJ, Khalaf A, Aizenstein HJ. Štúdium depresie pomocou zobrazovacích metód a metód strojového učenia.�NeuroImage Clin.�2016;10:115[Článok bez PMC]�[PubMed]
173.�Lanquillon S, Krieg JC, Bening-Abu-Shach U, Vedder H. Produkcia cytokínov a odpoveď na liečbu pri veľkej depresívnej poruche.�Neuropsychofarmak.�2000;22(4): 370 � 379.�[PubMed]
174.�Lindqvist D, Janelidze S, Erhardt S, Tr�skman-Bendz L, Engstr�m G, Brundin L. Biomarkery CSF pri pokusoch o samovraždu�analýza hlavnej zložky.�Acta Psychiatr Scand.�2011;124(1): 52 � 61.�[PubMed]
175.�Hidalgo-Mazzei D, Murru A, Reinares M, Vieta E, Colom F. Veľké údaje o duševnom zdraví: náročná fragmentovaná budúcnosť.�Svetová psychiatria.�2016;15(2): 186 � 187.�[Článok bez PMC]�[PubMed]
176.�Konzorcium C-DGotPG Identifikácia rizikových lokusov so spoločnými účinkami na päť hlavných psychiatrických porúch: analýza celého genómu.�Lancet.�2013;381(9875): 1371 � 1379.�[Článok bez PMC]�[PubMed]
177.�Dipnall JF, Pasco JA, Berk M, a kol. Spojenie dolovania údajov, strojového učenia a tradičných štatistík na detekciu biomarkerov spojených s depresiou.�PLoS One.�2016;11(2):e0148195.�[Článok bez PMC][PubMed]
178.�K�hler O, Benros ME, Nordentoft M, a kol. Účinok protizápalovej liečby na depresiu, depresívne symptómy a nežiaduce účinky: systematický prehľad a metaanalýza randomizovaných klinických štúdií.�Psychiatria JAMA.�2014;71(12): 1381 � 1391.�[PubMed]
179.�Wolkowitz OM, Reus VI, Chan T, a kol. Antiglukokortikoidná liečba depresie: dvojito zaslepený ketokonazol.�Biol Psychiatria.�1999;45(8): 1070 � 1074.�[PubMed]
180.�McAllister-Williams RH, Anderson IM, Finkelmeyer A, a kol. Antidepresívna augmentácia s metyrapónom pri depresii rezistentnej na liečbu (štúdia ADD): dvojito zaslepená, randomizovaná, placebom kontrolovaná štúdia.�Psychiatria Lancet.�2016;3(2): 117 � 127.�[PubMed]
181.�Gallagher P, Young AH. Mifepriston (RU-486) liečba depresie a psychózy: Prehľad terapeutických dôsledkov.�Neuropsychiatr Dis Treat.�2006;2(1): 33 � 42.�[Článok bez PMC]�[PubMed]
182.�Otte C, Hinkelmann K, Moritz S, a kol. Modulácia mineralokortikoidného receptora ako prídavná liečba pri depresii: randomizovaná, dvojito zaslepená, placebom kontrolovaná štúdia proof-of-concept.�J Psychiatr Res.�2010;44(6): 339 � 346.�[PubMed]
183.�Ozbolt LB, Nemeroff CB. Modulácia osi HPA pri liečbe porúch nálady.�Psychiatrická porucha.�2013;51:1147.
184.�Walker AK, Budac DP, Bisulco S, a kol. Blokáda NMDA receptora ketamínom ruší depresívne správanie vyvolané lipopolysacharidmi u myší C57BL/6J.�Neuropsychofarmak.�2013;38(9): 1609 � 1616.�[Článok bez PMC]�[PubMed]
185.�Lesp�rance F, Frasure-Smith N, St-Andr� E, Turecki G, Lesp�rance P, Wisniewski SR. Účinnosť suplementácie omega-3 pre veľkú depresiu: randomizovaná kontrolovaná štúdia.�Psychiatria J Clin.�2010;72(8): 1054 � 1062.�[PubMed]
186.�Kim S, Bae K, Kim J a kol. Použitie statínov na liečbu depresie u pacientov s akútnym koronárnym syndrómom.�Transl Psychiatria.�2015;5(8):e620.�[Článok bez PMC]�[PubMed]
187.�Shishehbor MH, Brennan ML, Aviles RJ a kol. Statíny podporujú silné systémové antioxidačné účinky prostredníctvom špecifických zápalových ciest.�Obeh.�2003;108(4): 426 � 431.�[PubMed]
188.�Mercier A, Auger-Aubin I, Lebeau JP a kol. Dôkaz o predpisovaní antidepresív na nepsychiatrické stavy v primárnej starostlivosti: analýza usmernení a systematické prehľady.�Rodinná prax BMC.�2013;14(1): 55.�[Článok bez PMC]�[PubMed]
189.�Freland L, Beaulieu JM. Inhibícia GSK3 lítiom, od jednotlivých molekúl po signálne siete.�Front Mol Neurosci.�2012;5:14[Článok bez PMC]�[PubMed]
190.�Horowitz MA, Zunszain PA. Neuroimunitné a neuroendokrinné abnormality pri depresii: dve strany tej istej mince.�Ann NY Acad Sci.�2015;1351(1): 68 � 79.�[PubMed]
191.�Juruena MF, Cleare AJ. Prekrytie medzi atypickou depresiou, sezónnou afektívnou poruchou a syndrómom chronickej únavy.�Rev Bras Psiquiatr.�2007;29:S19�S26.�[PubMed]
192.�Castr�n E, Kojima M. Neurotrofický faktor odvodený od mozgu pri poruchách nálady a liečbe antidepresívami.�Neurobiol Dis.�2017;97(Pt B): 119�126.�[PubMed]
193.�Pan A, Keum N, Okereke OI a kol. Obojsmerná súvislosť medzi depresiou a metabolickým syndrómom systematický prehľad a metaanalýza epidemiologických štúdií.�Diabetes Care.�2012;35(5): 1171 � 1180.�[Článok bez PMC]�[PubMed]
194.�Carvalho AF, Rocha DQ, McIntyre RS a kol. Adipokíny ako vznikajúce biomarkery depresie: systematický prehľad a metaanalýza.�J Psychiatric Res.�2014;59:28[PubMed]
195.�Wise T, Cleare AJ, Herane A, Young AH, Arnone D. Diagnostická a terapeutická užitočnosť neurozobrazovania pri depresii: prehľad.�Neuropsychiatr Dis Treat.�2014;10:1509.[Článok bez PMC]�[PubMed]
196.�Tamatam A, Khanum F, Bawa AS. Genetické biomarkery depresie.�Indián J Hum Genet.�2012;18(1): 20.�[Článok bez PMC]�[PubMed]
197.�Yoshimura R, Nakamura J, Shinkai K, Ueda N. Klinická odpoveď na liečbu antidepresívami a hladiny 3-metoxy-4-hydroxyfenylglykolu: mini prehľad.�Prog Neuropsychopharmacol Biol Psychiatry.�2004;28(4): 611 � 616.�[PubMed]
198.�Pierscionek T, Adekunte O, Watson S, Ferrier N, Alabi A. Úloha kortikosteroidov v antidepresívnej odpovedi.�ChronoPhys Ther.�2014;4:87.
199.�Poslanec Hage, Azar ST. Súvislosť medzi funkciou štítnej žľazy a depresiou.�J Thyroid Res.�2012;2012:590648[Článok bez PMC]�[PubMed]
200.�Dunn EC, Brown RC, Dai Y, a kol. Genetické determinanty depresie: nedávne zistenia a budúce smerovanie.�Harv Rev Psychiatry.�2015;23(1): 1.�[Článok bez PMC]�[PubMed]
201.�Yang CC, Hsu YL. Prehľad nositeľných detektorov pohybu na báze akcelerometrie na monitorovanie fyzickej aktivity.�Senzory.�2010;10(8): 7772 � 7788.�[Článok bez PMC]�[PubMed]
Artritída bolesť je komplexný jav zahŕňajúci zložité neurofyziologické spracovanie na všetkých úrovniach dráhy bolesti. Dostupné možnosti liečby na zmiernenie bolesti kĺbov sú dosť obmedzené a väčšina pacientov s artritídou uvádza len miernu úľavu od bolesti pri súčasnej liečbe. Lepšie pochopenie nervových mechanizmov zodpovedných za muskuloskeletálnu bolesť a identifikácia nových cieľov pomôže pri vývoji budúcich farmakologických terapií. Tento článok uvádza prehľad niektorých najnovších výskumov faktorov, ktoré prispievajú k bolestiam kĺbov a zahŕňa oblasti, ako sú kanabinoidy, receptory aktivované proteinázou, sodíkové kanály, cytokíny a prechodné kanály potenciálu receptorov. Diskutuje sa aj o vznikajúcej hypotéze, že osteoartritída môže mať neuropatickú zložku.
úvod
Svetová zdravotnícka organizácia zaraďuje ochorenia pohybového ústrojenstva ako najčastejšiu príčinu invalidity v modernom svete, ktorá postihuje každého tretieho dospelého [1]. Ešte alarmujúcejšie je, že prevalencia týchto chorôb narastá, zatiaľ čo naše poznatky o ich základných príčinách sú dosť základné.
Obr. 1 Schéma znázorňujúca niektoré z cieľov, o ktorých je známe, že modulujú bolesť kĺbov. Neuromodulátory sa môžu uvoľňovať z nervových zakončení, ako aj zo žírnych buniek a makrofágov, aby sa zmenila aferentná mechanosenzitivita. Endovaniloidy, kyselina a škodlivé teplo môžu aktivovať prechodný receptorový potenciál vaniloidných iónových kanálov typu 1 (TRPV1), čo vedie k uvoľneniu algogénnej substancie P (SP), ktorá sa následne viaže na receptory neurokinínu-1 (NK1). Proteázy môžu štiepiť a stimulovať proteázou aktivované receptory (PAR). Doteraz sa ukázalo, že PAR2 a PAR4 senzibilizujú primárne aferenty kĺbov. Endokanabinoidný anandamid (AE) sa vyrába na požiadanie a syntetizuje z N-arachidonoyl fosfatidyletanolamínu (NAPE) enzymatickým pôsobením fosfolipáz. Časť AE sa potom viaže na kanabinoidné-1 (CB1) receptory, čo vedie k neuronálnej desenzibilizácii. Nenaviazaný AE je rýchlo absorbovaný anandamidovým membránovým transportérom (AMT) predtým, než je štiepený amidhydrolázou mastnej kyseliny (FAAH) na etanolamín (Et) a kyselinu arachidónovú (AA). Cytokíny tumor nekrotizujúci faktor-a (TNF-a), interleukín-6 (IL-6) a interleukín1-beta (IL-1p) sa môžu viazať na svoje príslušné receptory, aby sa zvýšil prenos bolesti. Nakoniec, sodíkové kanály rezistentné na tetrodotoxín (TTX) (Nav1.8) sa podieľajú na senzibilizácii neurónov.
Pacienti túžia po svojom chronická bolesť zmiznúť; v súčasnosti predpisované analgetiká sú však väčšinou neúčinné a sú sprevádzané širokým spektrom nežiaducich vedľajších účinkov. Milióny ľudí na celom svete preto trpia vysiľujúcimi účinkami bolesti kĺbov, pre ktorú neexistuje uspokojivá liečba [2].
Viac ako 100 rôznych foriem artritídy má osteoartrózu (OA), ktorá je najbežnejšia. OA je progresívne degeneratívne ochorenie kĺbov, ktoré spôsobuje chronickú bolesť a stratu funkcie. Bežne je OA neschopnosťou kĺbu účinne opraviť poškodenie v reakcii na nadmerné sily, ktoré naň pôsobia. Biologické a psychosociálne faktory, ktoré zahŕňajú chronickú bolesť OA, nie sú dobre známe, hoci prebiehajúci výskum odhaľuje komplexnú povahu symptómov ochorenia [2]. Súčasné terapeutiká, ako sú nesteroidné protizápalové lieky (NSAID), poskytujú určitú symptomatickú úľavu, znižujú bolesť na krátke časové obdobia, ale nezmierňujú bolesť počas celého života pacienta. Okrem toho sa vysoké dávky NSAID nemôžu užívať opakovane počas mnohých rokov, pretože to môže viesť k renálnej toxicite a gastrointestinálnemu krvácaniu.
Tradične sa výskum artritídy zameriaval prevažne na kĺbovú chrupavku ako primárny cieľ pre terapeutický vývoj nových liekov s OA na modifikáciu ochorenia. Toto chondrogénne zameranie vrhlo nové svetlo na zložité biochemické a biomechanické faktory, ktoré ovplyvňujú správanie chondrocytov v chorých kĺboch. Keďže je však kĺbová chrupavka aneurálna a avaskulárna, je nepravdepodobné, že by toto tkanivo bolo zdrojom bolesti pri OA. Táto skutočnosť spolu so zistením, že neexistuje žiadna korelácia medzi poškodením kĺbovej chrupavky a bolesťou u pacientov s OA [3,4] alebo predklinickými modelmi OA [5], spôsobila posun v zameraní na vývoj liekov na účinnú kontrolu bolesti. . Tento článok zhodnotí najnovšie zistenia vo výskume bolesti kĺbov a upozorní na niektoré nové ciele, ktoré môžu byť budúcnosťou liečby bolesti pri artritíde (zhrnuté na obr. 1).
Cytokíny
Pôsobenie rôznych cytokínov v spoločných neurofyziologických štúdiách sa nedávno objavilo pomerne výrazne. Interleukín-6 (IL-6) je napríklad cytokín, ktorý sa typicky viaže na membránovo viazaný IL-6 receptor (IL-6R). IL-6 môže tiež signalizovať väzbou s rozpustným IL-6R (SIL-6R) za vzniku komplexu IL-6/sIL-6R. Tento komplex IL-6/sIL-6R sa následne viaže na transmembránovú glykoproteínovú podjednotku 130 (gp130), čím umožňuje IL-6 signalizovať v bunkách, ktoré konštitutívne neexprimujú membránovo viazaný IL-6R [25,26]. IL-6 a SIL-6R sú kľúčovými hráčmi pri systémovom zápale a artritíde, keďže v sére a synoviálnej tekutine pacientov s RA sa zistila zvýšená regulácia oboch [27,29]. Nedávno Vazquez et al. pozorovali, že súbežné podávanie IL-6/sIL-6R do kolien potkanov spôsobilo bolesť vyvolanú zápalom, ako sa ukázalo zvýšením odozvy neurónov chrbtového rohu chrbtice na mechanickú stimuláciu kolena a iných častí. zadnej končatiny [30]. Hyperexcitabilita miechových neurónov bola tiež pozorovaná, keď sa IL-6/sIL-6R aplikoval lokálne do miechy. Spinálna aplikácia rozpustného gp130 (ktorá by vyčistila komplexy IL-6/sIL-6R, čím by sa znížila trans-signalizácia) inhibovala centrálnu senzibilizáciu vyvolanú IL-6/sIL-6R. Akútna aplikácia samotného rozpustného gp130 však neznížila neuronálne reakcie na už vzniknutý zápal kĺbov.
Kanály prechodného receptorového potenciálu (TRP) sú neselektívne katiónové kanály, ktoré pôsobia ako integrátory rôznych fyziologických a patofyziologických procesov. Okrem termosenzácie, chemosenzácie a mechanosenzácie sa kanály TRP podieľajú na modulácii bolesti a zápalu. Napríklad sa ukázalo, že iónové kanály TRP vaniloid-1 (TRPV1) prispievajú k zápalovej bolesti kĺbov, keďže tepelná hyperalgézia nebola u TRPV1 monoartritických myší vyvolaná [31]. Podobne, TRP ankyrín-1 (TRPA1) iónové kanály sa podieľajú na artritickej mechano hypersenzitivite, pretože blokáda receptora selektívnymi antagonistami zmierňuje mechanickú bolesť vo Freundovom kompletnom adjuvantnom modeli zápalu [32,33]. Ďalší dôkaz, že TRPV1 sa môže podieľať na neurotransmisii bolesti OA, pochádza zo štúdií, v ktorých je expresia TRPV1 v neurónoch zvýšená v modeli OA s monojódacetátom sodným [34]. Okrem toho systémové podávanie antagonistu TRPV1 A-889425 znížilo evokovanú a spontánnu aktivitu miechového dynamického rozsahu a neurónov špecifických pre nocicepciu v monojódacetátovom modeli [35]. Tieto údaje naznačujú, že endovaniloidy by sa mohli podieľať na procesoch centrálnej senzibilizácie spojených s bolesťou OA.
V súčasnosti sú známe najmenej štyri polymorfizmy v géne, ktorý kóduje TRPV1, čo vedie k zmene štruktúry iónového kanála a zhoršenej funkcii. Jeden konkrétny polymorfizmus (rs8065080) mení citlivosť TRPV1 na kapsaicín a jedinci nesúci tento polymorfizmus sú menej citliví na tepelnú hyperalgéziu [36]. Nedávna štúdia skúmala, či pacienti s OA s polymorfizmom rs8065080 zaznamenali zmenené vnímanie bolesti na základe tejto genetickej anomálie. Výskumný tím zistil, že pacienti s asymptomatickou OA kolena mali väčšiu pravdepodobnosť, že budú nositeľmi génu rs8065080 ako pacienti s bolestivými kĺbmi. [37]. Toto pozorovanie naznačuje, že pacienti s OA s normálnym fungovaním; Kanály TRPV1 majú zvýšené riziko bolesti kĺbov a opätovne potvrdzujú potenciálne zapojenie TRPV1 do vnímania bolesti OA.
záver
Zatiaľ čo prekážka efektívneho liečenia bolesti pri artritíde zostáva, veľké skoky sa robia v našom chápaní neurofyziologických procesov zodpovedných za vznik bolesti kĺbov. Neustále sa objavujú nové ciele, zatiaľ čo mechanizmy za známymi dráhami sa ďalej definujú a zdokonaľujú. Je nepravdepodobné, že by riešením na normalizáciu bolesti kĺbov bolo zacielenie na jeden špecifický receptor alebo iónový kanál, ale skôr je indikovaný polyfarmaceutický prístup, pri ktorom sa počas špecifických fáz ochorenia používajú rôzne mediátory v kombinácii. Rozlúštenie funkčných obvodov na každej úrovni dráhy bolesti tiež zlepší naše znalosti o tom, ako bolesť kĺbov vzniká. Napríklad identifikácia periférnych mediátorov bolesti kĺbov nám umožní kontrolovať nocicepciu v kĺbe a pravdepodobne sa vyhnúť centrálnym vedľajším účinkom systémovo podávaných farmakoterapeutík.
FACETOGENICKÁ BOLESŤ
FACETOVÝ SYNDRÓM A FACETOGENICKÁ BOLESŤ
Facetov syndróm je kĺbová porucha súvisiaca s bedrovými fazetovými kĺbmi a ich inerváciami a spôsobuje lokálnu aj vyžarujúcu facetogénnu bolesť.
Nadmerná rotácia, extenzia alebo flexia chrbtice (opakované nadmerné zaťaženie) môže mať za následok degeneratívne zmeny kĺbovej chrupavky. Okrem toho môže zahŕňať degeneratívne zmeny iných štruktúr, vrátane medzistavcovej platničky.
SYNDRÓM KRČNEJ FACETY A FACETOGENICKÁ BOLESŤ
Axiálna bolesť krku (zriedkavo vyžarujúca cez ramená), najčastejšie jednostranná.
Bolesť s a/alebo obmedzením extenzie a rotácie
Citlivosť pri palpácii
Vyžarujúca facetogénna bolesť lokálne alebo do ramien alebo hornej časti chrbta a zriedkavo vyžarujúca do prednej alebo dolnej časti paže alebo do prstov ako herniovaný disk.
SYNDRÓM BEDROVEJ FACETY A FACETOGENICKÁ BOLESŤ
Bolesť alebo citlivosť v dolnej časti chrbta.
Miestna citlivosť / stuhnutosť pozdĺž chrbtice v dolnej časti chrbta.
Bolesť, stuhnutosť alebo ťažkosti s určitými pohybmi (ako je vzpriamené postavenie alebo vstávanie zo stoličky.
Bolesť pri hyperextenzii
Prenesená bolesť z horných bedrových fazetových kĺbov sa môže rozšíriť do boku, bedra a hornej laterálnej časti stehna.
Prenesená bolesť z dolných bedrových fazetových kĺbov môže preniknúť hlboko do stehna, laterálne a/alebo posteriórne.
Fazetové kĺby L4-L5 a L5-S1 môžu odkazovať na bolesť siahajúcu do distálnej laterálnej nohy a v zriedkavých prípadoch aj do chodidla
MEDICÍNA ZALOŽENÁ NA DÔKAZOCH
Intervenčná medicína proti bolesti založená na dôkazoch podľa klinických diagnóz
12. Bolesť pochádzajúca z bedrových fazetových kĺbov
abstraktné
Hoci existencia fazetového syndrómu bola dlho spochybňovaná, v súčasnosti je všeobecne akceptovaný ako klinická jednotka. V závislosti od diagnostických kritérií predstavujú zygapofyzické kĺby 5 % až 15 % prípadov chronickej axiálnej bolesti dolnej časti chrbta. Najčastejšie je facetogénna bolesť výsledkom opakovaného stresu a/alebo kumulatívnej traumy nízkej úrovne, čo vedie k zápalu a natiahnutiu kĺbového puzdra. Najčastejšou sťažnosťou je axiálna bolesť dolnej časti chrbta s odkazovanou bolesťou vnímanou v boku, bedre a stehne. Žiadne nálezy fyzikálneho vyšetrenia nie sú patognomické pre diagnózu. Najsilnejším indikátorom pre lumbálnu facetogénnu bolesť je zníženie bolesti po anestetických blokoch rami mediales (mediálnych vetiev) rami dorsales, ktoré inervujú fazetové kĺby. Pretože sa môžu vyskytnúť falošne pozitívne a možno aj falošne negatívne výsledky, výsledky sa musia interpretovať opatrne. U pacientov s injekčne potvrdenou zygapofyzickou bolesťou kĺbov možno vykonať procedurálne intervencie v kontexte multidisciplinárneho, multimodálneho liečebného režimu, ktorý zahŕňa farmakoterapiu, fyzikálnu terapiu a pravidelné cvičenie a, ak je to indikované, aj psychoterapiu. V súčasnosti je zlatým štandardom liečby facetogénnej bolesti rádiofrekvenčná liečba (1 B+). Dôkazy podporujúce intraartikulárne kortikosteroidy sú obmedzené; preto by to malo byť vyhradené pre tých, ktorí nereagujú na rádiofrekvenčnú liečbu (2 B1).
Facetogénna bolesť vychádzajúca z bedrových fazetových kĺbov je bežnou príčinou bolestí krížov u dospelej populácie. Goldthwaite bol prvý, kto opísal syndróm v roku 1911, a Ghormleymu sa všeobecne pripisuje vytvorenie termínu „fazetový syndróm“ v roku 1933. Facetogénna bolesť je definovaná ako bolesť, ktorá vzniká akoukoľvek štruktúrou, ktorá je súčasťou fazetových kĺbov, vrátane vláknitého puzdra. synoviálna membrána, hyalínna chrupavka a kosť.35
Častejšie je to výsledok opakovaného stresu a/alebo kumulatívnej traumy nízkej úrovne. To vedie k zápalu, ktorý môže spôsobiť naplnenie fazetového kĺbu tekutinou a opuch, čo vedie k natiahnutiu kĺbového puzdra a následnej tvorbe bolesti.27 Zápalové zmeny okolo fazetového kĺbu môžu tiež dráždiť miechový nerv prostredníctvom zúženia foraminálneho otvoru, čo vedie k ischias. Okrem toho Igarashi et al.28 zistili, že zápalové cytokíny uvoľňované cez ventrálne kĺbové puzdro u pacientov s degeneráciou zygapofyzálneho kĺbu môžu byť čiastočne zodpovedné za neuropatické symptómy u jedincov s spinálnou stenózou. Predisponujúce faktory pre zygapofyzickú bolesť kĺbov zahŕňajú spondylolistézu/lýzu, degeneratívne ochorenie disku a pokročilý vek.5
DODATOČNÉ SKÚŠKY IC
Miera prevalencie patologických zmien vo fazetových kĺboch pri rádiologickom vyšetrení závisí od priemerného veku subjektov, použitej rádiologickej techniky a definície abnormality. Degeneratívne fazetové kĺby možno najlepšie zobraziť pomocou vyšetrenia počítačovou tomografiou (CT).. 49
NEUROPATICKÁ BOLESŤ
Bolesť iniciovaná alebo spôsobená primárnou léziou alebo dysfunkciou v somatosenzorickom nervovom systéme.
Neuropatická bolesť je zvyčajne chronická, ťažko liečiteľná a často odolná voči štandardnej analgetickej liečbe.
abstraktné
Neuropatická bolesť je spôsobená léziou alebo chorobou somatosenzorického systému, vrátane periférnych vlákien (Ap, Ap a C vlákna) a centrálnych neurónov, a postihuje 7-10 % bežnej populácie. Bolo popísaných viacero príčin neuropatickej bolesti. Jeho výskyt sa pravdepodobne zvýši v dôsledku starnutia globálnej populácie, zvýšeného diabetes mellitus a zlepšeného prežívania rakoviny po chemoterapii. Nerovnováha medzi excitačnou a inhibičnou somatosenzorickou signalizáciou, zmeny v iónových kanáloch a variabilita v tom, ako sú správy o bolesti modulované v centrálnom nervovom systéme, sa podieľajú na neuropatickej bolesti. Okrem toho sa zdá, že bremeno chronickej neuropatickej bolesti súvisí so zložitosťou neuropatických symptómov, zlými výsledkami a ťažkými rozhodnutiami o liečbe. Dôležité je, že kvalita života je u pacientov s neuropatickou bolesťou znížená v dôsledku zvýšeného predpisovania liekov a návštev u poskytovateľov zdravotnej starostlivosti a chorobnosti zo samotnej bolesti a vyvolávajúcej choroby. Napriek výzvam pokrok v chápaní patofyziológie neuropatickej bolesti podnecuje vývoj nových diagnostických postupov a personalizovaných intervencií, ktoré zdôrazňujú potrebu multidisciplinárneho prístupu k manažmentu neuropatickej bolesti.
PATOGENÉZA NEUROPATICKEJ BOLESTI
PERIFÉRNE MECHANIZMY
Po lézii periférneho nervu sa neuróny stávajú citlivejšími a rozvíja sa u nich abnormálna excitabilita a zvýšená citlivosť na stimuláciu.
Toto je známe ako... Periférna senzibilizácia!
CENTRÁLNE MECHANIZMY
V dôsledku prebiehajúcej spontánnej aktivity na periférii sa v neurónoch vyvíja zvýšená aktivita pozadia, zväčšené receptívne polia a zvýšené reakcie na aferentné impulzy, vrátane normálnych hmatových podnetov. Toto je známe ako...Centrálna senzibilizácia!
Chronická neuropatická bolesť je častejšia u žien (8 % oproti 5.7 % u mužov) a u pacientov vo veku > 50 rokov (8.9 % oproti 5.6 % u mladších ako 49 rokov) a najčastejšie postihuje dolnú časť chrbta a dolné končatiny , krk a horné končatiny24. Lumbálne a krčné bolestivé radikulopatie sú pravdepodobne najčastejšou príčinou chronickej neuropatickej bolesti. V súlade s týmito údajmi prieskum viac ako 12,000 40 pacientov s chronickou bolesťou s nociceptívnymi aj neuropatickými typmi bolesti, postúpený odborníkom na bolesť v Nemecku, odhalil, že 25 % všetkých pacientov pociťovalo aspoň niektoré charakteristiky neuropatickej bolesti (ako sú pocity pálenia, necitlivosť a brnenie); postihnutí boli najmä pacienti s chronickou bolesťou chrbta a radikulopatiouXNUMX.
Príspevok klinickej neurofyziológie k pochopeniu mechanizmov tenznej bolesti hlavy.
abstraktné
Doteraz sa uskutočnili klinické neurofyziologické štúdie tenznej bolesti hlavy (TTH) s dvoma hlavnými cieľmi: (1) zistiť, či niektoré neurofyziologické parametre môžu pôsobiť ako markery TTH, a (2) preskúmať fyziopatológiu TTH. Pokiaľ ide o prvý bod, súčasné výsledky sú sklamaním, pretože niektoré abnormality zistené u pacientov s TTH môžu byť často pozorované aj u migrenikov. Na druhej strane klinická neurofyziológia zohrala dôležitú úlohu v diskusii o patogenéze TTH. Štúdie o exteroceptívnom potlačení kontrakcie temporalis svalu odhalili dysfunkciu excitability mozgového kmeňa a suprasegmentálnej kontroly. K podobnému záveru sa dospelo použitím trigeminocervikálnych reflexov, ktorých abnormality v TTH naznačujú zníženú inhibičnú aktivitu interneurónov mozgového kmeňa, čo odráža abnormálne endogénne mechanizmy kontroly bolesti. Je zaujímavé, že abnormalita nervovej excitability v TTH sa zdá byť všeobecným javom, ktorý sa neobmedzuje len na kraniálne oblasti. Defektné mechanizmy podobné DNIC boli skutočne dokázané aj v somatických oblastiach štúdiami nociceptívnych flexných reflexov. Bohužiaľ, väčšina neurofyziologických štúdií o TTH je poznačená vážnymi metodologickými chybami, ktorým by sa malo v budúcom výskume vyhnúť, aby sa lepšie objasnili mechanizmy TTH.
Referencie:
Neurofyziológia bolesti pri artritíde. McDougall JJ1 Linton P.
Bolesť pochádzajúca z bedrových fazetových kĺbov. van Kleef M1,Vanelderen P,Cohen SP,Lataster A,Van Zundert J,Mekhail N.
Neuropatická bolesťLuana Colloca,1Taylor Ludman,1Didier Bouhassira,2Ralf Baron,3Anthony H. Dickenson,4David Yarnitsky,5Roy Freeman,6Andrea Truiniová,7Nadine Attalová, Nanna B. Finnerup,9Christopher Eccleston,10,11Eija Kalso,12David L. Bennett,13Robert H. Dworkin,14a Srinivasa N. Raja15
Príspevok klinickej neurofyziológie k pochopeniu mechanizmov tenznej bolesti hlavy. Rossi P1, Vollono C, Valeriani M, Sandrini G.
Lekári definujú chronickú bolesť ako akúkoľvek bolesť, ktorá trvá 3 až 6 mesiacov alebo dlhšie. The bolesť ovplyvňuje duševné zdravie jednotlivca a každodenný život. Bolesť pochádza zo série správ, ktoré prechádzajú nervovým systémom. Zdá sa, že depresia nasleduje po bolesti. Spôsobuje vážne symptómy, ktoré ovplyvňujú to, ako sa jednotlivec cíti, myslí a ako zvláda každodenné činnosti, tj spánok, jedenie a prácu. Chiropraktik, Dr. Alex Jimenez, sa ponorí do potenciálnych biomarkerov, ktoré môžu pomôcť pri hľadaní a liečbe základných príčin bolesti a chronickej bolesti.
Prvým krokom k úspešnej liečbe bolesti je komplexné biopsychosociálne posúdenie.
Rozsah organickej patológie sa nemusí presne odrážať v prežívaní bolesti.
Počiatočné hodnotenie možno použiť na identifikáciu oblastí, ktoré si vyžadujú hlbšie hodnotenie.
Na posúdenie vplyvu chronickej bolesti je k dispozícii mnoho overených nástrojov na sebareportáž.
Hodnotenie pacientov s chronickou bolesťou
Chronická bolesť je problémom verejného zdravia, ktorý postihuje 20 – 30 % populácie západných krajín. Hoci došlo k mnohým vedeckým pokrokom v chápaní neurofyziológie bolesti, presné posúdenie a diagnostika pacientovho problému s chronickou bolesťou nie je priamočiare ani dobre definované. Spôsob, akým je chronická bolesť koncipovaná, ovplyvňuje spôsob hodnotenia bolesti a faktory, ktoré sa berú do úvahy pri stanovení diagnózy chronickej bolesti. Neexistuje žiadny priamy vzťah medzi množstvom alebo typom organickej patológie a intenzitou bolesti, ale namiesto toho, prežívanie chronickej bolesti je formované nespočetnými biomedicínskymi, psychosociálnymi (napr. presvedčenia, očakávania a nálada pacientov) a behaviorálnymi faktormi (napr. kontext, reakcie významných iných). Posúdenie každej z týchto troch domén prostredníctvom komplexného hodnotenia osoby s chronickou bolesťou je nevyhnutné pre rozhodnutia o liečbe a uľahčenie optimálnych výsledkov. Toto vyšetrenie by malo zahŕňať dôkladnú anamnézu pacienta a lekárske vyšetrenie a krátky skríningový rozhovor, na ktorom možno pozorovať správanie pacienta. Ďalšie hodnotenie na riešenie otázok identifikovaných počas počiatočného hodnotenia povedie k rozhodnutiam o tom, aké dodatočné hodnotenia, ak nejaké, môžu byť vhodné. K dispozícii sú štandardizované samostatne hlásené nástroje na vyhodnotenie intenzity bolesti pacienta, funkčných schopností, presvedčení a očakávaní a emocionálneho utrpenia, ktoré môže poskytnúť lekár, alebo môže poskytnúť odporúčanie na hĺbkové vyhodnotenie, ktoré pomôže pri plánovaní liečby.
Bolesť je mimoriadne prevládajúci príznak. Odhaduje sa, že samotná chronická bolesť postihuje 30 % dospelej populácie USA, viac ako 100 miliónov dospelých.1
Napriek stúpajúcim nákladom na liečbu ľudí s chronickou bolesťou je úľava pre mnohých stále nepolapiteľná a úplné odstránenie bolesti je zriedkavé. Hoci došlo k značnému pokroku v znalostiach neurofyziológie bolesti spolu s vývojom účinných analgetických liekov a iných inovatívnych lekárskych a chirurgických zákrokov, v priemere je miera zníženia bolesti dostupnými postupmi 30 – 40 % a to sa vyskytuje v menej ako polovica liečených pacientov.
Spôsob, akým premýšľame o bolesti, ovplyvňuje spôsob, akým ideme hodnotiť bolesť. Hodnotenie začína anamnézou a fyzikálnym vyšetrením, po ktorom nasledujú laboratórne testy a diagnostické zobrazovacie postupy v snahe identifikovať a/alebo potvrdiť prítomnosť akejkoľvek základnej patológie spôsobujúcej symptóm/y alebo generátor bolesti.
Pri absencii identifikovateľnej organickej patológie môže poskytovateľ zdravotnej starostlivosti predpokladať, že hlásenie symptómov pochádza z psychologických faktorov a môže požiadať o psychologické vyšetrenie na zistenie emocionálnych faktorov, ktoré sú základom pacientovej správy. Existuje dualita, kde sa hlásenie symptómov pripisuje jednému z nich somatická or psychogénne mechanizmy.
Napríklad organické základy pre niektoré z najbežnejších a opakujúcich sa akútnych (napr. bolesť hlavy)3 a chronických [napr. bolesť v chrbte, fibromyalgia (FM)] problémy s bolesťou sú do značnej miery neznáme,4,5 kým na druhej strane, asymptomatickí jedinci môžu mať štrukturálne abnormality, ako sú herniované platničky, ktoré by vysvetľovali bolesť, ak by bola prítomná.6,7�Chýbajú adekvátne vysvetlenia pre pacientov bez identifikovanej organickej patológie, ktorí uvádzajú silnú bolesť a bezbolestných jedincov s významnou objektívnou patológiou.
Chronická bolesť postihuje nielen samotného pacienta, ale aj jeho blízkych (partnerov, príbuzných, zamestnávateľov, spolupracovníkov a priateľov).), pričom je nevyhnutná vhodná liečba. Uspokojivá liečba môže pochádzať iba z komplexného posúdenia biologickej etiológie bolesti v spojení so špecifickou psychosociálnou a behaviorálnou prezentáciou pacienta, vrátane jeho emocionálneho stavu (napr. úzkosť, depresia a hnev), vnímania a chápania symptómov a reakcií na ne. symptómy významnými ostatnými.8,9 Kľúčovým predpokladom je, že symptómy a funkčné obmedzenia jedincov s chronickou bolesťou ovplyvňujú viaceré faktory. Preto je potrebné komplexné hodnotenie, ktoré sa bude zaoberať biomedicínskymi, psychosociálnymi a behaviorálnymi oblasťami, pretože každá z nich prispieva k chronickej bolesti a súvisiacemu postihnutiu.10,11
Komplexné hodnotenie jednotlivca s chronickou bolesťou
Turk a Meichenbaum12 navrhli, že hodnotenie ľudí, ktorí hlásia bolesť, by mali viesť tri hlavné otázky:
Aký je rozsah pacientovho ochorenia alebo úrazu (fyzického postihnutia)?
Aký je rozsah choroby? To znamená, do akej miery pacient trpí, je invalidný a nemôže sa tešiť z bežných aktivít?
Zdá sa, že správanie jednotlivca je primerané chorobe alebo zraneniu, alebo existujú dôkazy o zosilnení symptómov z rôznych psychologických alebo sociálnych dôvodov (napr. výhody ako pozitívna pozornosť, lieky na zmenu nálady, finančná kompenzácia)?
Na zodpovedanie týchto otázok je potrebné získať informácie od pacienta na základe anamnézy a fyzikálneho vyšetrenia v kombinácii s klinickým rozhovorom a prostredníctvom štandardizovaných hodnotiacich nástrojov. Poskytovatelia zdravotnej starostlivosti musia prostredníctvom fyzikálneho vyšetrenia a diagnostických testov hľadať akúkoľvek príčinu bolesti a súčasne hodnotiť pacientovu náladu, obavy, očakávania, úsilie o zvládanie, zdroje, reakcie významných druhých a vplyv bolesti na pacientov� žije.11 Stručne povedané, poskytovateľ zdravotnej starostlivosti musí hodnotiť „celého človeka“ a nielen bolesť.
Všeobecné ciele anamnézy a lekárskeho hodnotenia sú:
(ii) určiť, či lekárske údaje môžu vysvetliť symptómy pacienta, závažnosť symptómov a funkčné obmedzenia
iii) stanoviť lekársku diagnózu
iv) zhodnotiť dostupnosť vhodnej liečby
v) stanoviť ciele liečby
vi) určiť vhodný postup na zvládnutie symptómov, ak úplné vyliečenie nie je možné.
Značný počet pacientov, ktorí hlásia chronickú bolesť, nepreukázali žiadnu fyzickú patológiu pomocou jednoduchých röntgenových snímok, počítačovej axiálnej tomografie alebo elektromyografie (k dispozícii je rozsiahla literatúra o fyzickom hodnotení, rádiografických a laboratórnych hodnotiacich postupoch na určenie fyzického základu bolesti),17 čo sťažuje alebo znemožňuje presnú patologickú diagnózu.
Napriek týmto obmedzeniam zostáva anamnéza a fyzikálne vyšetrenie pacienta základom lekárskej diagnózy, môže poskytnúť ochranu proti nadmernej interpretácii zistení z diagnostického zobrazovania, ktoré je do značnej miery potvrdzujúce, a môže sa použiť na usmernenie ďalšieho úsilia v oblasti hodnotenia.
Okrem toho pacienti s problémami chronickej bolesti často konzumujú rôzne lieky.18 Počas rozhovoru je dôležité prediskutovať pacientove súčasné lieky, pretože mnohé lieky proti bolesti sú spojené s vedľajšími účinkami, ktoré môžu spôsobiť alebo napodobňovať emocionálny stres.19 Poskytovatelia zdravotnej starostlivosti by nemali poznať len lieky používané na chronickú bolesť, ale aj vedľajšie účinky týchto liekov, ktoré vedú k únave, ťažkostiam so spánkom a zmenám nálady, aby sa predišlo nesprávnej diagnóze depresie.
Používanie denných denníkov sa považuje za presnejšie, pretože sú založené na reálnom čase a nie na pamäti. Pacienti môžu byť požiadaní, aby si viedli pravidelné denníky intenzity bolesti s hodnoteniami zaznamenanými niekoľkokrát každý deň (napr. jedlo a spánok) počas niekoľkých dní alebo týždňov a viaceré hodnotenia bolesti možno spriemerovať v priebehu času.
Jeden problém zaznamenaný pri používaní papierových a ceruzkových denníkov je ten, že pacienti nemusia dodržiavať pokyny na poskytovanie hodnotenia v určených intervaloch. Pacienti môžu skôr vyplniť denníky vopred (�vyplniť dopredu�) alebo krátko pred návštevou lekára (�vyplniť dozadu�),24 čo podkopáva predpokladanú platnosť denníkov. Elektronické denníky si získali uznanie v niektorých výskumných štúdiách, aby sa predišlo týmto problémom.
Výskum preukázal dôležitosť hodnotenia celkovej kvality života súvisiacej so zdravím (HRQOL) u pacientov s chronickou bolesťou okrem funkcie.31,32 Existuje množstvo dobre zavedených, psychometricky podporovaných opatrení HRQOL [Medical Outcomes Study Short-Form Health Survey (SF-36)],33 všeobecné merania fyzického fungovania [napr. index bolestivých porúch (PDI)],34 a opatrenia špecifické pre chorobu [napr. MacMasterov index osteoartritídy Western Ontario (WOMAC);35 Roland-Morris Back Pain Disability Questionnaire (RDQ) )]36 na posúdenie funkcie a kvality života.
Opatrenia špecifické pre chorobu sú navrhnuté tak, aby zhodnotili vplyv špecifického stavu (napr. bolesť a stuhnutosť u ľudí s osteoartritídou), zatiaľ čo všeobecné opatrenia umožňujú porovnať fyzické fungovanie spojené s danou poruchou a jej liečbou s rôznymi inými stavmi. Pri použití generického opatrenia nemusia byť zistené špecifické účinky poruchy; preto opatrenia špecifické pre ochorenie môžu s väčšou pravdepodobnosťou odhaliť klinicky významné zlepšenie alebo zhoršenie špecifických funkcií v dôsledku liečby. Všeobecné merania fungovania môžu byť užitočné na porovnanie pacientov s rôznymi bolestivými stavmi. Kombinované používanie opatrení špecifických pre chorobu a generických opatrení uľahčuje dosiahnutie oboch cieľov.
Prítomnosť emocionálneho stresu u ľudí s chronickou bolesťou predstavuje problém pri hodnotení symptómov, ako je únava, znížená úroveň aktivity, znížené libido, zmena chuti do jedla, poruchy spánku, priberanie alebo strata hmotnosti a poruchy pamäti a koncentrácie, pretože tieto symptómy môžu byť výsledkom bolesti, emocionálneho stresu alebo liečebných liekov predpísaných na kontrolu bolesti.
Nástroje boli vyvinuté špeciálne pre pacientov trpiacich bolesťou, aby zhodnotili psychické utrpenie, vplyv bolesti na životy pacientov, pocit kontroly, zvládacie správanie a postoje k chorobe, bolesti a poskytovateľom zdravotnej starostlivosti.17
Napríklad Beck Depression Inventory (BDI)39 a Profil stavov nálady (POMS)40 sú psychometricky vhodné na hodnotenie symptómov depresívnej nálady, emočného stresu a porúch nálady a odporúča sa ich použitie vo všetkých klinických štúdiách chronická bolesť;41 však skóre sa musí interpretovať opatrne a môže byť potrebné upraviť kritériá pre úrovne emočného stresu, aby sa predišlo falošným pozitívam.42
Laboratórne biomarkery pre bolesť
Biomarkery sú biologické charakteristiky, ktoré možno použiť na označenie zdravia alebo choroby. Tento článok skúma štúdie o biomarkeroch bolesti dolnej časti chrbta (LBP) u ľudských subjektov. LBP je hlavnou príčinou invalidity spôsobenej rôznymi poruchami súvisiacimi s chrbticou, vrátane degenerácie medzistavcovej platničky, herniácie platničky, spinálnej stenózy a fazetovej artritídy. Ťažiskom týchto štúdií sú zápalové mediátory, pretože zápal prispieva k patogenéze degenerácie disku a súvisiacich mechanizmov bolesti. Štúdie čoraz častejšie naznačujú, že prítomnosť zápalových mediátorov možno merať systémovo v krvi. Tieto biomarkery môžu slúžiť ako nové nástroje na riadenie starostlivosti o pacienta. V súčasnosti je odpoveď pacienta na liečbu nepredvídateľná s významnou mierou recidívy, a hoci chirurgická liečba môže poskytnúť anatomickú korekciu a úľavu od bolesti, je invazívna a nákladná. Prehľad zahŕňa štúdie uskutočnené na populáciách so špecifickými diagnózami a nedefinovaným pôvodom LBP. Keďže prirodzená história LBP je progresívna, dočasný charakter štúdií je kategorizovaný podľa trvania symptómov/ochorenia. Súvisiace štúdie o zmenách biomarkerov s liečbou sú tiež preskúmané. V konečnom dôsledku majú diagnostické biomarkery LBP a degenerácie chrbtice potenciál viesť éru individualizovanej medicíny chrbtice pre personalizované terapeutiká pri liečbe LBP.
Biomarkery pre chronickú neuropatickú bolesť a potenciálnu aplikáciu pri stimulácii miechy
Tento prehľad bol zameraný na pochopenie toho, ktoré látky v ľudskom tele sa zvyšujú a znižujú so zvyšujúcou sa neuropatickou bolesťou. Preskúmali sme rôzne štúdie a videli sme korelácie medzi neuropatickou bolesťou a zložkami imunitného systému (tento systém chráni telo pred chorobami a infekciami). Naše zistenia budú užitočné najmä pre pochopenie spôsobov, ako znížiť alebo odstrániť nepohodlie, ktoré so sebou prináša chronická neuropatická bolesť. Postup stimulácie miechy (SCS) je jedným z mála pomerne účinných liečebných postupov na liečbu bolesti. Následná štúdia použije naše zistenia z tohto prehľadu na SCS, aby sme pochopili mechanizmus a ďalej optimalizovali účinnosť.
Zistilo sa, že prozápalové cytokíny, ako je IL-1p, IL-6, IL-2, IL-33, CCL3, CXCL1, CCR5 a TNF-a, hrajú významnú úlohu pri amplifikácii stavov chronickej bolesti.
Po preskúmaní rôznych štúdií týkajúcich sa biomarkerov bolesti sme zistili, že sérové hladiny prozápalových cytokínov a chemokínov, ako sú IL-1p, IL-6, IL-2, IL-33, CCL3, CXCL1, CCR5 a TNF -?, boli významne upregulované počas zážitku chronickej bolesti. Na druhej strane sa zistilo, že protizápalové cytokíny ako IL-10 a IL-4 vykazujú významnú down-reguláciu počas stavu chronickej bolesti.
Biomarkery pre depresiu
Množstvo výskumov zahŕňalo stovky predpokladaných biomarkerov depresie, ale ešte úplne neobjasnilo ich úlohu pri depresívnom ochorení ani nezistilo, čo je abnormálne u ktorých pacientov a ako možno biologické informácie použiť na zlepšenie diagnózy, liečby a prognózy. Tento nedostatočný pokrok je čiastočne spôsobený povahou a heterogenitou depresie v spojení s metodologickou heterogenitou v rámci výskumnej literatúry a veľkým množstvom biomarkerov s potenciálom, ktorých vyjadrenie sa často mení v závislosti od mnohých faktorov. Preskúmali sme dostupnú literatúru, ktorá naznačuje, že markery zapojené do zápalových, neurotrofických a metabolických procesov, ako aj zložky neurotransmiterov a neuroendokrinných systémov, predstavujú vysoko sľubných kandidátov. Tieto môžu byť merané prostredníctvom genetických a epigenetických, transkriptomických a proteomických, metabolomických a neuroimagingových hodnotení. Teraz sa vyžaduje použitie nových prístupov a systematických výskumných programov, aby sa určilo, či a ktoré biomarkery možno použiť na predpovedanie odpovede na liečbu, stratifikovať pacientov na špecifickú liečbu a vypracovať ciele pre nové intervencie. Dospeli sme k záveru, že existuje veľa prísľubov na zníženie bremena depresie prostredníctvom ďalšieho rozvoja a rozširovania týchto výskumných ciest.
Referencie:
Vyšetrenie pacientov s chronickou bolesťou�EJ Dansiet a DC Turk*t�
Zápalové biomarkery bolesti dolnej časti chrbta a degenerácie disku: prehľad.
Khan AN1, Jacobsen HE2, Khan J1, Filippi CG3, Levine M3, Lehman RA Jr2,4, Riew KD2,4, Lenke LG2,4, Chahine NO2,5.
Biomarkery pre chronickú neuropatickú bolesť a ich potenciálna aplikácia pri stimulácii miechy: prehľad
Chibueze D. Nwagwu, 1 Christina Sarris, MD, 3 Yuan-Xiang Tao, Ph.D., MD, 2 a Antonios Mammis, MD1,2
Biomarkery pre depresiu: nedávne poznatky, súčasné výzvy a vyhliadky do budúcnosti. Strawbridge R1, Young AH1,2, Cleare AJ1,2.
Bolesť je prirodzenou reakciou ľudského tela na zranenie alebo chorobu a často je varovaním, že niečo nie je v poriadku. Keď sa problém vylieči, vo všeobecnosti prestaneme pociťovať tieto bolestivé symptómy, čo sa však stane, keď bolesť pretrváva dlho potom, čo príčina pominie? Chronická bolesť je medicínsky definovaná ako pretrvávajúca bolesť, ktorá trvá 3 až 6 mesiacov alebo viac. Chronická bolesť je určite náročný stav, s ktorým sa dá žiť, ovplyvňuje všetko od úrovne aktivity jednotlivca a jeho schopnosti pracovať, ako aj jeho osobné vzťahy a psychologické podmienky. Uvedomujete si však, že chronická bolesť môže tiež ovplyvniť štruktúru a funkciu vášho mozgu? Ukazuje sa, že tieto zmeny mozgu môžu viesť k kognitívnym aj psychickým poruchám.
Chronická bolesť neovplyvňuje len jedinú oblasť mysle, v skutočnosti môže viesť k zmenám v mnohých základných oblastiach mozgu, z ktorých väčšina sa podieľa na mnohých základných procesoch a funkciách. Rôzne výskumné štúdie v priebehu rokov zistili zmeny hipokampu spolu so znížením šedej hmoty z dorzolaterálnej prefrontálnej kôry, amygdaly, mozgového kmeňa a pravej ostrovnej kôry, aby sme vymenovali aspoň niektoré, spojené s chronickou bolesťou. Rozdelenie niekoľkých štruktúr týchto oblastí a ich súvisiacich funkcií môže pomôcť dať tieto mozgové zmeny do kontextu u mnohých jedincov s chronickou bolesťou. Účelom nasledujúceho článku je demonštrovať a diskutovať o štrukturálnych a funkčných zmenách mozgu spojených s chronickou bolesťou, najmä v prípade, keď tieto pravdepodobne neodrážajú ani poškodenie, ani atrofiu.
Štrukturálne zmeny mozgu pri chronickej bolesti pravdepodobne neodrážajú ani poškodenie, ani atrofiu
abstraktné
Zdá sa, že chronická bolesť je spojená s redukciou šedej hmoty mozgu v oblastiach, ktoré možno pripísať prenosu bolesti. Morfologické procesy, ktoré sú základom týchto štrukturálnych zmien, pravdepodobne po funkčnej reorganizácii a centrálnej plasticite v mozgu, zostávajú nejasné. Bolesť pri koxartróze je jedným z mála chronických bolestivých syndrómov, ktoré sú v zásade liečiteľné. Vyšetrili sme 20 pacientov s chronickou bolesťou v dôsledku jednostrannej koxartrózy (priemerný vek 63.25 – 9.46 (SD) rokov, 10 žien) pred endoprotetickou operáciou bedrového kĺbu (bolestivý stav) a sledovali sme štrukturálne zmeny mozgu do 1 roka po operácii: 6 – 8 týždňov , 12�18 týždňov a 10�14 mesiacov, keď je úplne bez bolesti. Pacienti s chronickou bolesťou v dôsledku jednostrannej koxartrózy mali signifikantne menej šedej hmoty v porovnaní s kontrolami v prednom cingulárnom kortexe (ACC), ostrovčekovom kortexe a operkule, dorzolaterálnom prefrontálnom kortexe (DLPFC) a orbitofrontálnom kortexe. Tieto oblasti fungujú ako multiintegračné štruktúry počas prežívania a predvídania bolesti. Keď boli pacienti po zotavení z endoprotetickej operácie bez bolesti, zistilo sa zvýšenie šedej hmoty v takmer rovnakých oblastiach. Zistili sme tiež progresívny nárast šedej hmoty mozgu v premotorickej kôre a doplnkovej motorickej oblasti (SMA). Dospeli sme k záveru, že abnormality sivej hmoty pri chronickej bolesti nie sú príčinou, ale sú sekundárne k ochoreniu a sú aspoň čiastočne spôsobené zmenami v motorickej funkcii a telesnej integrácii.
úvod
Dôkazy o funkčnej a štrukturálnej reorganizácii u pacientov s chronickou bolesťou podporujú myšlienku, že chronická bolesť by nemala byť chápaná len ako zmenený funkčný stav, ale aj ako dôsledok funkčnej a štrukturálnej plasticity mozgu [1], [2], [3], [4], [5], [6]. Za posledných šesť rokov bolo publikovaných viac ako 20 štúdií demonštrujúcich štrukturálne zmeny mozgu pri 14 syndrómoch chronickej bolesti. Pozoruhodným rysom všetkých týchto štúdií je skutočnosť, že zmeny šedej hmoty neboli rozdelené náhodne, ale vyskytujú sa v definovaných a funkčne vysoko špecifických oblastiach mozgu – konkrétne zapojenie do supraspinálneho nociceptívneho spracovania. Najvýraznejšie nálezy boli odlišné pre každý bolestivý syndróm, ale prekrývali sa v cingulárnom kortexe, orbitofrontálnom kortexe, inzule a dorzálnom moste [4]. Ďalšie štruktúry zahŕňajú talamus, dorzolaterálny prefrontálny kortex, bazálne gangliá a hipokampálnu oblasť. Tieto nálezy sú často diskutované ako bunková atrofia, posilňujúca myšlienku poškodenia alebo straty sivej hmoty mozgu [7], [8], [9]. V skutočnosti vedci zistili koreláciu medzi poklesom sivej hmoty mozgu a trvaním bolesti [6], [10]. Trvanie bolesti je však tiež spojené s vekom pacienta a vekovo závislý globálny, ale aj regionálne špecifický pokles šedej hmoty je dobre zdokumentovaný [11]. Na druhej strane, týmito štrukturálnymi zmenami môže byť aj zmenšenie veľkosti buniek, extracelulárnych tekutín, synaptogenéza, angiogenéza alebo dokonca zmeny objemu krvi [4], [12], [13]. Nech je zdroj akýkoľvek, pre našu interpretáciu takýchto zistení je dôležité vidieť tieto morfometrické nálezy vo svetle množstva morfometrických štúdií plasticity závislej od cvičenia, vzhľadom na to, že po kognitívnom a fyzickom cvičení sa opakovane ukázali regionálne špecifické štrukturálne zmeny mozgu [ 14].
Nie je pochopiteľné, prečo sa syndróm chronickej bolesti rozvinie len u relatívne malej časti ľudí, keďže bolesť je univerzálna skúsenosť. Vzniká otázka, či u niektorých ľudí môže štrukturálny rozdiel v centrálnych systémoch prenášajúcich bolesť pôsobiť ako diatéza chronickej bolesti. Zmeny šedej hmoty pri fantómovej bolesti v dôsledku amputácie [15] a poranenia miechy [3] naznačujú, že morfologické zmeny mozgu sú aspoň čiastočne dôsledkom chronickej bolesti. Bolesť pri koxartróze (OA) je však jedným z mála syndrómov chronickej bolesti, ktorý je v zásade liečiteľný, pretože 88 % týchto pacientov pravidelne netrpí bolesťou po operácii totálnej náhrady bedrového kĺbu (THR) [16]. V pilotnej štúdii sme analyzovali desať pacientov s OA bedrového kĺbu pred a krátko po operácii. Zistili sme úbytok sivej hmoty v prednej cingulovanej kôre (ACC) a insule počas chronickej bolesti pred operáciou THR a zistili sme zvýšenie šedej hmoty v príslušných oblastiach mozgu v bezbolestnom stave po operácii [17]. So zameraním na tento výsledok sme teraz rozšírili naše štúdie skúmajúce viac pacientov (n?=?20) po úspešnej THR a monitorovali štrukturálne zmeny mozgu v štyroch časových intervaloch, až do jedného roka po operácii. Na kontrolu zmien šedej hmoty v dôsledku zlepšenia motoriky alebo depresie sme tiež poskytli dotazníky zamerané na zlepšenie motorických funkcií a duševného zdravia.
Materiály a metódy
Dobrovoľníci
Pacienti, ktorí sú tu uvedení, sú podskupinou 20 pacientov z 32 nedávno publikovaných pacientov, ktorí boli porovnaní so zdravou kontrolnou skupinou zodpovedajúceho veku a pohlavia [17], ale zúčastnili sa ďalšieho jednoročného sledovania. Po chirurgickom zákroku 12 pacientov odstúpilo z dôvodu druhej endoprotetickej operácie (n=~2), ťažkého ochorenia (n~=~2) a odvolania súhlasu (n~=~8). Takto zostala skupina dvadsiatich pacientov s jednostrannou primárnou OA bedrového kĺbu (priemerný vek 63.25 – 9.46 (SD) rokov, 10 žien), ktorí boli vyšetrovaní štyrikrát: pred operáciou (bolestivý stav) a znova 6, 8 a 12, 18 týždňov a 10 �14 mesiacov po endoprotetickej operácii, keď je úplne bez bolesti. Všetci pacienti s primárnou OA bedrového kĺbu mali anamnézu bolesti dlhšiu ako 12 mesiacov v rozsahu od 1 do 33 rokov (priemer 7.35 roka) a priemerné skóre bolesti 65.5 (v rozsahu 40 až 90) na vizuálnej analógovej škále (VAS) v rozsahu od 0 (žiadna bolesť) až 100 (najhoršia predstaviteľná bolesť). Hodnotili sme akýkoľvek výskyt menších bolestivých udalostí, vrátane zubov, uší a bolesti hlavy až 4 týždne pred štúdiou. Náhodne sme vybrali aj údaje z 20 zdravých kontrol so zhodným pohlavím a vekom (priemerný vek 60,95 – 8,52 (SD) rokov, 10 žien) z 32 z vyššie uvedenej pilotnej štúdie [17]. Žiadny z 20 pacientov alebo z 20 zdravých dobrovoľníkov rovnakého pohlavia a veku nemal žiadnu neurologickú alebo internú anamnézu. Štúdia bola eticky schválená miestnou etickou komisiou a pred vyšetrením bol získaný písomný informovaný súhlas od všetkých účastníkov štúdie.
Údaje o správaní
Údaje o depresii, somatizácii, úzkosti, bolesti a fyzickom a duševnom zdraví u všetkých pacientov a vo všetkých štyroch časových bodoch sme zbierali pomocou nasledujúcich štandardizovaných dotazníkov: Beck Depression Inventory (BDI) [18], Brief Symptom Inventory (BSI) [19], Schmerzempfindungs-Skala (SES?=?škála nepríjemnosti bolesti) [20] a 36-položkový krátky formulár zdravotného prieskumu (SF-36) [21] a Nottinghamský zdravotný profil (NHP). Uskutočnili sme opakované merania ANOVA a spárovali sme obojstranné t-testy na analýzu dlhodobých behaviorálnych údajov pomocou SPSS 13.0 pre Windows (SPSS Inc., Chicago, IL) a použili sme korekciu Greenhouse Geisser, ak bol porušený predpoklad sféricity. Hladina významnosti bola stanovená na p<0.05.
VBM – Data Acquisition
Zber obrázkov. MR skenovanie s vysokým rozlíšením sa uskutočnilo na 3T MRI systéme (Siemens Trio) so štandardnou 12-kanálovou hlavovou cievkou. Pre každý zo štyroch časových bodov sken I (medzi 1 dňom a 3 mesiacmi pred endoprotetickou operáciou), sken II (6 až 8 týždňov po operácii), sken III (12 až 18 týždňov po operácii) a sken IV (10�14 mesiacov po operácii) sa pre každého pacienta získala T1 vážená štrukturálna MRI pomocou 3D-FLASH sekvencie (TR 15 ms, TE 4.9 ms, uhol otočenia 25�, 1 mm rezy, FOV 256�256, veľkosť voxelu 1�1� 1 mm).
Spracovanie obrazu a štatistická analýza
Predspracovanie a analýza údajov sa uskutočňovali pomocou SPM2 (Wellcome Department of Cognitive Neurology, Londýn, Veľká Británia) bežiaceho pod Matlab (Mathworks, Sherborn, MA, USA) a obsahujúceho voxelovú morfometriu (VBM)-toolbox pre pozdĺžne údaje, ktoré je založený na štrukturálnych 3D MR obrázkoch s vysokým rozlíšením a umožňuje použiť voxelovú štatistiku na detekciu regionálnych rozdielov v hustote alebo objemoch šedej hmoty [22], [23]. Stručne povedané, predbežné spracovanie zahŕňalo priestorovú normalizáciu, segmentáciu šedej hmoty a 10 mm priestorové vyhladzovanie pomocou gaussovského jadra. Pre kroky predspracovania sme použili optimalizovaný protokol [22], [23] a šablónu šedej hmoty špecifickú pre skener a štúdiu [17]. Použili sme skôr SPM2 ako SPM5 alebo SPM8, aby bola táto analýza porovnateľná s našou pilotnou štúdiou [17]. pretože umožňuje výbornú normalizáciu a segmentáciu longitudinálnych dát. Keďže však bola nedávno k dispozícii novšia aktualizácia VBM (VBM8) (dbm.neuro.uni-jena.de/vbm/), použili sme aj VBM8.
Prierezová analýza
Použili sme dvojvzorkový t-test, aby sme zistili regionálne rozdiely v sivej hmote mozgu medzi skupinami (pacienti v časovom bode skenovania I (chronická bolesť) a zdravé kontroly). Použili sme prah p<0.001 (nekorigovaný) na celý mozog kvôli našej silnej apriornej hypotéze, ktorá je založená na 9 nezávislých štúdiách a kohortách preukazujúcich pokles šedej hmoty u pacientov s chronickou bolesťou [7], [8], [ 9], [15], [24], [25], [26], [27], [28], že nárast šedej hmoty sa objaví v rovnakých oblastiach (relevantných pre spracovanie bolesti) ako v našej pilotnej štúdii (17 ). Skupiny boli spárované podľa veku a pohlavia bez významných rozdielov medzi skupinami. Aby sme zistili, či sa rozdiely medzi skupinami zmenili po jednom roku, porovnali sme pacientov aj pri IV skenovaní časového bodu (bez bolesti, jednoročné sledovanie) s našou zdravou kontrolnou skupinou.
Pozdĺžna analýza
Na zistenie rozdielov medzi časovými bodmi (skenovanie I�IV) sme porovnávali skeny pred operáciou (stav bolesti) a znova 6�8 a 12�18 týždňov a 10�14 mesiacov po endoprotetickej operácii (bez bolesti) ako ANOVA s opakovaným meraním. Pretože akékoľvek zmeny na mozgu v dôsledku chronickej bolesti môžu potrebovať určitý čas na ústup po operácii a vymiznutí bolesti a kvôli bolestiam po operácii, ktoré pacienti hlásili, porovnávali sme v longitudinálnej analýze sken I a II so skenom III a IV. Na zistenie zmien, ktoré nie sú úzko spojené s bolesťou, sme tiež hľadali progresívne zmeny vo všetkých časových intervaloch. Prevrátili sme mozgy pacientov s OA ľavého bedra (n?=?7), aby sme sa normalizovali na stranu bolesti pre skupinové porovnanie aj pozdĺžnu analýzu, ale primárne sme analyzovali neprevrátené údaje. V modeli sme použili skóre BDI ako kovariát.
výsledky
Behaviorálne údaje
Všetci pacienti hlásili chronickú bolesť bedrového kĺbu pred operáciou a boli bez bolesti (pokiaľ ide o túto chronickú bolesť) bezprostredne po chirurgickom zákroku, ale hlásili skôr akútnu bolesť po chirurgickom zákroku na skene II, ktorá sa líšila od bolesti spôsobenej osteoartritídou. Skóre duševného zdravia SF-36 (F(1.925/17.322)?=?0.352, p?=?0.7) a globálne skóre BSI GSI (F(1.706/27.302)?=?3.189, p?=?0.064 ) nevykazovali žiadne zmeny v priebehu času a žiadnu duševnú komorbiditu. Žiadna z kontrolných skupín neuviedla akútnu alebo chronickú bolesť a žiadna nevykazovala žiadne príznaky depresie alebo fyzického/mentálneho postihnutia.
Pred operáciou niektorí pacienti vykazovali mierne až stredne závažné depresívne symptómy v skóre BDI, ktoré sa významne znížilo na skenovaní III (t(17)?=?2.317, p?=?0.033) a IV (t(16)?=?2.132, p? = 0.049). Okrem toho sa skóre SES (nepríjemnosť bolesti) všetkých pacientov výrazne zlepšilo od skenu I (pred operáciou) po sken II (t(16)?=~4.676, p<0.001), sken III (t(14)?=? 4.760, p<0.001) a sken IV (t(14)?=?4.981, p<0.001, 1 rok po operácii), pretože nepríjemnosť bolesti klesala s intenzitou bolesti. Hodnotenie bolesti na skene 1 a 2 bolo pozitívne, rovnaké hodnotenie na 3. a 4. deň negatívne. SES popisuje iba kvalitu vnímanej bolesti. Preto bol pozitívny na 1. a 2. deň (priemer 19.6 na 1. a 13.5 na 2. deň) a negatívny (na) na 3. a 4. deň. Niektorí pacienti však tomuto postupu nerozumeli a použili SES ako globálnu kvalitu životnej miery. To je dôvod, prečo boli všetci pacienti v ten istý deň individuálne a tou istou osobou požiadaní o výskyt bolesti.
V krátkom prieskume zdravia (SF-36), ktorý pozostáva zo súhrnných meraní skóre fyzického zdravia a skóre duševného zdravia [29], sa pacienti výrazne zlepšili v skóre fyzického zdravia od skenu I po sken II (t( 17) ?=?? 4.266, p = ? 0.001), sken III (t(16) ? = 8.584, p < 0.001) a IV (t(12) ? = 7.148, p < 0.001), ale nie v skóre duševného zdravia. Výsledky NHP boli podobné, v subškále �bolesť� (obrátená polarita) sme pozorovali významnú zmenu zo skenu I na sken II (t(14)?=??5.674, p<0.001, sken III (t(12) )?=??7.040, p<0.001 a sken IV (t(10)?=??3.258, p?=?0.009). Zistili sme tiež významné zvýšenie subškály „fyzická mobilita“ od skenu I po sken III (t(12)?=??3.974, p?=?0.002) a sken IV (t(10)?=??2.511, p?=?0.031). Medzi skenom I a skenom II nebola žiadna významná zmena ( šesť týždňov po operácii).
Štrukturálne údaje
Prierezová analýza. Do všeobecného lineárneho modelu sme zahrnuli vek ako kovariát a nezistili sme žiadne vekové zmätky. V porovnaní s kontrolami zhodnými s pohlavím a vekom pacienti s primárnou OA bedrového kĺbu (n?=~20) vykazovali predoperačne (Scan I) zníženú šedú hmotu v prednom cingulárnom kortexe (ACC), ostrovčekovom kortexe, operkule, dorzolaterálnom prefrontálnom kortexe ( DLPFC), pravý temporálny pól a cerebellum (tabuľka 1 a obrázok 1). Okrem pravého putamenu (x?=?31, y?=??14, z?=??1; p<0.001, t?=?3.32) nebol u pacientov s OA zistený signifikantný nárast hustoty šedej hmoty v porovnaní s k zdravým kontrolám. Porovnaním pacientov v časovom bode skenu IV so zodpovedajúcimi kontrolami sa zistili rovnaké výsledky ako v prierezovej analýze s použitím skenu I v porovnaní s kontrolami.
Obrázok 1: Štatistické parametrické mapy demonštrujúce štrukturálne rozdiely v sivej hmote u pacientov s chronickou bolesťou v dôsledku primárnej OA bedrového kĺbu v porovnaní s kontrolami a pozdĺžne v porovnaní so sebou samými v priebehu času. Významné zmeny šedej hmoty sú zobrazené farebne superponované, údaje v priereze sú znázornené červenou a pozdĺžne údaje sú znázornené žltou farbou. Axiálna rovina: ľavá strana obrázku je ľavá strana mozgu. hore: Oblasti významného poklesu sivej hmoty medzi pacientmi s chronickou bolesťou v dôsledku primárnej OA bedrového kĺbu a nepostihnutými kontrolnými subjektmi. p<0.001 nekorigované dno: Nárast sivej hmoty u 20 pacientov bez bolesti v treťom a štvrtom období skenovania po operácii totálnej náhrady bedrového kĺbu v porovnaní s prvým (predoperačným) a druhým (6-8 týždňov po operácii) skenom. p<0.001 nekorigované Grafy: Odhady kontrastu a 90% interval spoľahlivosti, efekty záujmu, ľubovoľné jednotky. os x: kontrasty pre 4 časové body, os y: odhad kontrastu pri ?3, 50, 2 pre ACC a odhad kontrastu pri 36, 39, 3 pre ostrovček.
Prevrátením údajov pacientov s OA ľavého bedra (n?=?7) a ich porovnaním so zdravými kontrolami sa výsledky významne nezmenili, avšak pre pokles talamu (x?=?10, y?=??20, z?=?3, p<0.001, t?=?3.44) a zväčšenie pravého mozočku (x?=?25, y?=??37, z?=??50, p<0.001, t? =>5.12), ktoré nedosiahli významnosť v neprevrátených údajoch pacientov v porovnaní s kontrolami.
Pozdĺžna analýza. Pri longitudinálnej analýze sa zistil významný nárast (p < 001 nekorigovaný) sivej hmoty pri porovnaní prvého a druhého skenovania (chronická bolesť/pooperačná bolesť) s tretím a štvrtým skenom (bez bolesti) v ACC, ostrovčekovej kôry, cerebellum a pars orbitalis u pacientov s OA (tabuľka 2 a obrázok 1). Šedá hmota sa časom zmenšila (p < 001 nekorigovaná analýza celého mozgu) v sekundárnej somatosenzorickej kôre, hipokampe, strednom cingulárnom kortexe, talame a jadre kaudátu u pacientov s OA (obrázok 2).
Obrázok 2: a) Významné zvýšenie sivej hmoty mozgu po úspešnej operácii. Axiálny pohľad na významný pokles šedej hmoty u pacientov s chronickou bolesťou v dôsledku primárnej OA bedrového kĺbu v porovnaní s kontrolnými subjektmi. p<0.001 nekorigované (prierezová analýza), b) Pozdĺžne zvýšenie šedej hmoty v priebehu času v žltom porovnávacom skene I&IIscan III>scan IV) u pacientov s OA. p<0.001 nekorigované (pozdĺžna analýza). Ľavá strana obrázku je ľavá strana mozgu.
Prevrátením údajov pacientov s OA ľavého bedra (n?=?7) sa výsledky významne nezmenili, ale pre zníženie šedej hmoty mozgu v Heschlovom gyrus (x?=??41, y?=?? 21, z?=?10, p<0.001, t=?3.69) a Precuneus (x?=?15, y?=??36, z?=?3, p<0.001, t?=?4.60) .
Porovnaním prvého skenu (predoperačný) so skenmi 3+4 (pooperačný) sme zistili nárast šedej hmoty vo frontálnom kortexe a motorickom kortexe (p<0.001 nekorigované). Všimli sme si, že tento kontrast je menej prísny, pretože teraz máme menej skenov na jeden stav (bolesť vs. bez bolesti). Keď prah znížime, zopakujeme to, čo sme zistili pomocou kontrastu 1+2 vs. 3+4.
Hľadaním oblastí, ktoré sa zväčšujú vo všetkých časových intervaloch, sme zistili zmeny šedej hmoty mozgu v motorických oblastiach (oblasť 6) u pacientov s koxartrózou po totálnej náhrade bedrového kĺbu (sken Idbm.neuro.uni-jena.de/vbm/) by sme mohli replikovať tento nález v prednej a strednej cingulárnej kôre a oboch predných insulae.
Vypočítali sme veľkosti účinkov a prierezová analýza (pacienti vs. kontroly) poskytla Cohen�sd 1.78751 v píku voxel ACC (x?=??12, y?=?25, z?=?? 16). Vypočítali sme aj Cohen�sd pre pozdĺžnu analýzu (kontrastný sken 1+2 vs. sken 3+4). Výsledkom bol Cohen�sd 1.1158 v ACC (x?=??3, y?=?50, z?=?2). Pokiaľ ide o ostrovček (x?=?33, y?=?21, z?=?13) a vo vzťahu k rovnakému kontrastu, Cohen�sd je 1.0949. Okrem toho sme vypočítali priemer nenulových hodnôt voxelov mapy Cohen�sd v rámci ROI (pozostávajúcej z prednej časti gyrus cingulate a subcallosal cortex, odvodenej z Harvard-Oxford Cortical Structural Atlas): 1.251223.
Pohľad Dr. Alexa Jimeneza
Pacienti s chronickou bolesťou môžu v priebehu času pociťovať rôzne zdravotné problémy, okrem ich už tak oslabujúcich symptómov. Napríklad veľa jedincov bude mať v dôsledku bolesti problémy so spánkom, ale čo je najdôležitejšie, chronická bolesť môže viesť aj k rôznym problémom duševného zdravia, vrátane úzkosti a depresie. Účinky, ktoré môže mať bolesť na mozog, sa môžu zdať až príliš ohromujúce, ale stále viac dôkazov naznačuje, že tieto zmeny mozgu nie sú trvalé a možno ich zvrátiť, keď pacienti s chronickou bolesťou dostanú správnu liečbu svojich základných zdravotných problémov. Podľa článku abnormality sivej hmoty zistené pri chronickej bolesti neodrážajú poškodenie mozgu, ale sú skôr reverzibilným dôsledkom, ktorý sa normalizuje, keď je bolesť adekvátne liečená. Našťastie sú dostupné rôzne liečebné prístupy, ktoré pomáhajú zmierniť symptómy chronickej bolesti a obnoviť štruktúru a funkciu mozgu.
Diskusia
Monitorovaním celej štruktúry mozgu v priebehu času potvrdzujeme a rozširujeme naše nedávno publikované pilotné údaje [17]. Zistili sme zmeny v sivej hmote mozgu u pacientov s primárnou koxartrózou v stave chronickej bolesti, ktoré sa čiastočne zvrátia, keď sú títo pacienti bez bolesti, po endoprotetickej operácii bedrového kĺbu. Čiastočný nárast šedej hmoty po operácii je takmer v tých istých oblastiach, kde bol pred operáciou pozorovaný pokles šedej hmoty. Prevrátenie údajov pacientov s OA ľavého bedra (a teda normalizácia na stranu bolesti) malo len malý vplyv na výsledky, ale navyše ukázalo zníženie šedej hmoty v Heschlovom gyrus a Precuneus, ktoré nevieme ľahko vysvetliť a, keďže neexistuje žiadna apriórna hypotéza, berte to s veľkou opatrnosťou. Avšak rozdiel medzi pacientmi a zdravými kontrolami pri skenovaní I bol stále pozorovateľný v prierezovej analýze pri skenovaní IV. Relatívny nárast šedej hmoty v priebehu času je preto jemný, tj nie dostatočne zreteľný, aby mal vplyv na analýzu prierezu, čo už bolo preukázané v štúdiách skúmajúcich plasticitu závislú od skúseností [30], [31]. Poznamenávame, že skutočnosť, že ukazujeme, že niektoré časti zmien mozgu v dôsledku chronickej bolesti sú reverzibilné, nevylučuje, že niektoré ďalšie časti týchto zmien sú nezvratné.
Je zaujímavé, že sme pozorovali, že pokles šedej hmoty v ACC u pacientov s chronickou bolesťou pred operáciou zrejme pokračuje 6 týždňov po operácii (skenovanie II) a zvyšuje sa iba smerom k skenovaniu III a IV, pravdepodobne v dôsledku bolesti po operácii alebo zníženia motoriky. funkciu. To je v súlade s behaviorálnymi údajmi skóre fyzickej mobility zahrnuté v NHP, ktoré pooperačne nevykazovalo žiadnu významnú zmenu v časovom bode II, ale významne sa zvýšilo smerom k skenovaniu III a IV. Je potrebné poznamenať, že naši pacienti po operácii neuviedli žiadnu bolesť v bedre, ale pociťovali pooperačnú bolesť v okolitých svaloch a koži, ktorú pacienti vnímali veľmi odlišne. Keďže však pacienti stále hlásili určitú bolesť pri skenovaní II, porovnali sme aj prvé skenovanie (pred chirurgickým zákrokom) so skenmi III + IV (po operácii), čo odhalilo nárast šedej hmoty vo frontálnom kortexe a motorickom kortexe. Všimli sme si, že tento kontrast je menej prísny z dôvodu menšieho počtu skenov na stav (bolesť vs. bez bolesti). Keď sme znížili prah, zopakujeme to, čo sme zistili pomocou kontrastu I+II vs. III+IV.
Naše údaje silne naznačujú, že zmeny šedej hmoty u pacientov s chronickou bolesťou, ktoré sa zvyčajne nachádzajú v oblastiach zapojených do supraspinálneho nociceptívneho spracovania [4], nie sú spôsobené atrofiou neurónov ani poškodením mozgu. Skutočnosť, že tieto zmeny pozorované v stave chronickej bolesti sa úplne nezvrátia, možno vysvetliť relatívne krátkym obdobím pozorovania (jeden rok po operácii oproti priemeru siedmich rokov chronickej bolesti pred operáciou). Neuroplastické zmeny mozgu, ktoré sa mohli vyvinúť počas niekoľkých rokov (ako dôsledok neustáleho nociceptívneho vstupu), pravdepodobne potrebujú viac času na úplné zvrátenie. Ďalšou možnosťou, prečo je možné nárast sivej hmoty detegovať iba v pozdĺžnych údajoch, ale nie v údajoch v priereze (tj medzi kohortami v časovom bode IV), je, že počet pacientov (n?=~20) je príliš malý. Je potrebné zdôrazniť, že rozptyl medzi mozgami niekoľkých jedincov je dosť veľký a že pozdĺžne údaje majú tú výhodu, že rozptyl je relatívne malý, keďže tie isté mozgy sa skenujú niekoľkokrát. V dôsledku toho budú jemné zmeny zistiteľné iba v longitudinálnych údajoch [30], [31], [32]. Samozrejme nemôžeme vylúčiť, že tieto zmeny sú aspoň čiastočne ireverzibilné, aj keď je to nepravdepodobné, vzhľadom na zistenia špecifickej štrukturálnej plasticity a reorganizácie [4], [12], [30], [33], [34]. Na zodpovedanie tejto otázky musia budúce štúdie opakovane vyšetrovať pacientov počas dlhších časových období, možno rokov.
Poznamenávame, že môžeme urobiť len obmedzené závery týkajúce sa dynamiky morfologických zmien mozgu v priebehu času. Dôvodom je to, že keď sme navrhli túto štúdiu v roku 2007 a skenovali v rokoch 2008 a 2009, nebolo známe, či vôbec nastanú štrukturálne zmeny a z dôvodov uskutočniteľnosti sme zvolili dátumy a časové rámce skenovania, ako je tu opísané. Dalo by sa tvrdiť, že k zmenám sivej hmoty v čase, ktoré popisujeme pre skupinu pacientov, mohlo dôjsť aj v kontrolnej skupine (časový efekt). Akékoľvek zmeny v dôsledku starnutia, ak vôbec, by sa však dali očakávať ako zníženie objemu. Vzhľadom na našu apriórnu hypotézu založenú na 9 nezávislých štúdiách a kohortách preukazujúcich pokles šedej hmoty u pacientov s chronickou bolesťou [7], [8], [9], [15], [24], [25], [26], [27], [28] sme sa zamerali na regionálne nárasty v čase, a preto veríme, že naše zistenie nie je jednoduchým časovým efektom. Je potrebné poznamenať, že nemôžeme vylúčiť, že pokles šedej hmoty v priebehu času, ktorý sme našli v našej skupine pacientov, by mohol byť spôsobený časovým účinkom, pretože sme našu kontrolnú skupinu neskenovali v rovnakom časovom rámci. Vzhľadom na tieto zistenia by sa budúce štúdie mali zamerať na viac a kratšie časové intervaly, keďže morfometrické zmeny mozgu závislé od cvičenia sa môžu vyskytnúť tak rýchlo ako po 1 týždni [32], [33].
Okrem vplyvu nociceptívneho aspektu bolesti na šedú hmotu mozgu [17], [34] sme pozorovali, že k štrukturálnym zmenám pravdepodobne prispievajú aj zmeny v motorickej funkcii. Zistili sme, že motorické a premotorické oblasti (oblasť 6) sa zvyšujú vo všetkých časových intervaloch (obrázok 3). Intuitívne to môže byť spôsobené zlepšením motorických funkcií v priebehu času, pretože pacienti už neboli obmedzovaní v bežnom živote. Predovšetkým sme sa nezamerali na motorickú funkciu, ale na zlepšenie prežívania bolesti, vzhľadom na našu pôvodnú snahu zistiť, či je dobre známe zníženie sivej hmoty mozgu u pacientov s chronickou bolesťou v zásade reverzibilné. V dôsledku toho sme nepoužili špecifické nástroje na skúmanie motorickej funkcie. Napriek tomu je (funkčná) reorganizácia motorickej kôry u pacientov s bolestivými syndrómami dobre zdokumentovaná [35], [36], [37], [38]. Okrem toho je motorická kôra jedným z cieľov v terapeutických prístupoch u pacientov s ťažko zvládnuteľnou chronickou bolesťou pomocou priamej mozgovej stimulácie [39], [40], transkraniálnej stimulácie jednosmerným prúdom [41] a opakovanej transkraniálnej magnetickej stimulácie [42], [43]. Presné mechanizmy takejto modulácie (facilitácia vs. inhibícia alebo jednoducho interferencia v sieťach súvisiacich s bolesťou) ešte nie sú objasnené [40]. Nedávna štúdia preukázala, že špecifický motorický zážitok môže zmeniť štruktúru mozgu [13]. Synaptogenéza, reorganizácia pohybových reprezentácií a angiogenéza v motorickej kôre sa môžu vyskytnúť so špeciálnymi požiadavkami na motorickú úlohu. Tsao a spol. preukázali reorganizáciu v motorickej kôre pacientov s chronickou bolesťou dolnej časti chrbta, ktorá sa zdá byť špecifická pre bolesť chrbta [44] a Puri et al. pozorovali redukciu šedej hmoty ľavej doplnkovej motorickej oblasti u pacientov s fibromyalgiou [45]. Naša štúdia nebola navrhnutá tak, aby rozlúštila rôzne faktory, ktoré môžu zmeniť mozog pri chronickej bolesti, ale naše údaje týkajúce sa zmien šedej hmoty interpretujeme tak, že neodrážajú výlučne dôsledky neustáleho nociceptívneho vstupu. V skutočnosti nedávna štúdia u pacientov s neuropatickou bolesťou poukázala na abnormality v oblastiach mozgu, ktoré zahŕňajú emocionálne, autonómne a vnímanie bolesti, čo naznačuje, že hrajú rozhodujúcu úlohu v globálnom klinickom obraze chronickej bolesti [28].
Obrázok 3: Štatistické parametrické mapy preukazujúce významný nárast šedej hmoty mozgu v motorických oblastiach (oblasť 6) u pacientov s koxartrózou pred a po THR (pozdĺžna analýza, sken I Odhady kontrastu pri x?=?19, y?=??12, z?=?70.
Dve nedávne pilotné štúdie sa zamerali na terapiu náhrady bedrového kĺbu u pacientov s osteoartrózou, čo je jediný syndróm chronickej bolesti, ktorý je v zásade liečiteľný totálnou náhradou bedrového kĺbu [17], [46] a tieto údaje dopĺňa veľmi nedávna štúdia u pacientov s chronickou bolesťou dolnej časti chrbta [ 47]. Tieto štúdie je potrebné vnímať vo svetle niekoľkých longitudinálnych štúdií skúmajúcich neuronálnu plasticitu závislú od skúseností u ľudí na štrukturálnej úrovni [30], [31] a nedávnej štúdie o štrukturálnych zmenách mozgu u zdravých dobrovoľníkov, ktorí zažívali opakovanú bolestivú stimuláciu [34]. . Kľúčovým posolstvom všetkých týchto štúdií je, že hlavný rozdiel v štruktúre mozgu medzi pacientmi s bolesťou a kontrolnou skupinou môže ustúpiť, keď sa bolesť vylieči. Je však potrebné vziať do úvahy, že jednoducho nie je jasné, či sú zmeny u pacientov s chronickou bolesťou spôsobené výlučne nociceptívnym vstupom alebo následkami bolesti alebo oboma spôsobmi. Je viac ako pravdepodobné, že zmeny správania, ako je deprivácia alebo posilnenie sociálnych kontaktov, agilita, telesný tréning a zmeny životného štýlu sú dostatočné na formovanie mozgu [6], [12], [28], [48]. Najmä depresia ako komorbidita alebo dôsledok bolesti je kľúčovým kandidátom na vysvetlenie rozdielov medzi pacientmi a kontrolami. Malá skupina našich pacientov s OA vykazovala mierne až stredne závažné depresívne symptómy, ktoré sa časom menili. Nezistili sme, že štrukturálne zmeny sa výrazne zhodujú so skóre BDI, ale vyvstáva otázka, koľko ďalších zmien správania v dôsledku absencie bolesti a zlepšenia motoriky môže prispieť k výsledkom a do akej miery. Tieto zmeny správania môžu ovplyvniť zníženie šedej hmoty pri chronickej bolesti, ako aj zvýšenie šedej hmoty, keď bolesť zmizne.
Ďalším dôležitým faktorom, ktorý môže skresľovať našu interpretáciu výsledkov, je skutočnosť, že takmer všetci pacienti s chronickou bolesťou užívali lieky proti bolesti, ktoré vysadili, keď už boli bez bolesti. Dalo by sa tvrdiť, že NSAID ako diklofenak alebo ibuprofén majú určité účinky na nervový systém a to isté platí pre opioidy, antiepileptiká a antidepresíva, lieky, ktoré sa často používajú pri liečbe chronickej bolesti. Vplyv liekov proti bolesti a iných liekov na morfometrické nálezy môže byť dôležitý (48). Žiadna štúdia doteraz nepreukázala účinky liekov proti bolesti na morfológiu mozgu, ale niekoľko prác zistilo, že zmeny v štruktúre mozgu u pacientov s chronickou bolesťou nie sú vysvetlené iba nečinnosťou súvisiacou s bolesťou [15], ani liekmi proti bolesti [7], [9], [49]. Chýbajú však konkrétne štúdie. Ďalší výskum by sa mal zamerať na zmeny v kortikálnej plasticite závislé od skúseností, ktoré môžu mať rozsiahle klinické dôsledky na liečbu chronickej bolesti.
Zistili sme tiež pokles šedej hmoty v pozdĺžnej analýze, pravdepodobne v dôsledku reorganizačných procesov, ktoré sprevádzajú zmeny v motorickej funkcii a vnímaní bolesti. Je málo dostupných informácií o pozdĺžnych zmenách sivej hmoty mozgu pri bolestivých stavoch, z tohto dôvodu nemáme žiadnu hypotézu o poklese sivej hmoty v týchto oblastiach po operácii. Teutsch a kol. [25] zistili zvýšenie sivej hmoty mozgu v somatosenzorickom a strednom cingulárnom kortexe u zdravých dobrovoľníkov, ktorí pociťovali bolestivú stimuláciu v dennom protokole počas ôsmich po sebe nasledujúcich dní. Zistenie zvýšenia sivej hmoty po experimentálnom nociceptívnom vstupe sa anatomicky do určitej miery prekrývalo s poklesom sivej hmoty mozgu v tejto štúdii u pacientov, ktorí boli vyliečení z dlhotrvajúcej chronickej bolesti. To znamená, že nociceptívny vstup u zdravých dobrovoľníkov vedie k štrukturálnym zmenám závislým od cvičenia, ako je to možné u pacientov s chronickou bolesťou, a že tieto zmeny sa u zdravých dobrovoľníkov zvrátia, keď nociceptívny vstup prestane. V dôsledku toho možno pokles šedej hmoty v týchto oblastiach pozorovaný u pacientov s OA interpretovať tak, že sleduje rovnaký základný proces: zmeny mozgu závislé od cvičenia [50]. Ako neinvazívna procedúra je MR Morfometria ideálnym nástrojom na hľadanie morfologických substrátov chorôb, prehĺbenie nášho chápania vzťahu medzi štruktúrou a funkciou mozgu a dokonca aj na sledovanie terapeutických zásahov. Jednou z veľkých výziev v budúcnosti je prispôsobiť tento výkonný nástroj pre multicentrické a terapeutické štúdie chronickej bolesti.
Obmedzenia tejto štúdie
Hoci táto štúdia je rozšírením našej predchádzajúcej štúdie, ktorá rozširuje údaje o sledovaní na 12 mesiacov a skúma viac pacientov, naše zásadné zistenie, že morfometrické zmeny mozgu pri chronickej bolesti sú reverzibilné, je dosť jemné. Veľkosti účinkov sú malé (pozri vyššie) a účinky sú čiastočne poháňané ďalšou redukciou regionálneho objemu mozgovej šedej hmoty v časovom bode skenovania 2. Keď vylúčime údaje zo skenu 2 (priamo po operácii), iba významné zvýšenie sivej hmoty mozgu pre motorickú kôru a prednú kôru prežije prah p < 0.001 nekorigovaný (tabuľka 3).
záver
Nie je možné rozlíšiť, do akej miery sú štrukturálne zmeny, ktoré sme pozorovali, spôsobené zmenami nocicepčného vstupu, zmenami v motorickej funkcii alebo spotrebou liekov alebo zmenami v blahobyte ako takom. Maskovanie skupinových kontrastov prvého a posledného skenovania medzi sebou odhalilo oveľa menej rozdielov, ako sa očakávalo. Zmeny mozgu spôsobené chronickou bolesťou so všetkými dôsledkami sa pravdepodobne vyvíjajú pomerne dlho a môžu tiež potrebovať nejaký čas, aby sa vrátili. Napriek tomu tieto výsledky odhaľujú procesy reorganizácie, čo silne naznačuje, že chronický nociceptívny vstup a motorické poškodenie u týchto pacientov vedie k zmenenému spracovaniu v kortikálnych oblastiach a následne k štrukturálnym zmenám mozgu, ktoré sú v princípe reverzibilné.
Poďakovanie
Ďakujeme všetkým dobrovoľníkom za účasť na tejto štúdii a skupine Fyzika a metódy v NeuroImage Nord v Hamburgu. Štúdia bola eticky schválená miestnou etickou komisiou a pred vyšetrením bol získaný písomný informovaný súhlas od všetkých účastníkov štúdie.
Vyhlásenie o financovaní
Táto práca bola podporená grantmi DFG (Nemecká výskumná nadácia) (MA 1862/2-3) a BMBF (Spolkové ministerstvo školstva a výskumu) (371 57 01 a NeuroImage Nord). Investori nezohrávali žiadnu úlohu pri navrhovaní štúdie, zbere a analýze údajov, rozhodovaní o publikovaní alebo príprave rukopisu.
Endokanabinoidný systém: Základný systém, o ktorom ste nikdy nepočuli
V prípade, že ste ešte nepočuli o endokanabinoidnom systéme alebo ECS, nemusíte sa hanbiť. V 1960. rokoch 30. storočia vyšetrovatelia, ktorí sa začali zaujímať o biologickú aktivitu kanabisu, nakoniec izolovali mnohé z jeho aktívnych chemikálií. Trvalo však ďalších XNUMX rokov, kým výskumníci študujúci zvieracie modely našli receptor pre tieto chemikálie ECS v mozgu hlodavcov, čo je objav, ktorý otvoril celý svet skúmania existencie receptorov ECS a ich fyziologického účelu.
Teraz vieme, že väčšina zvierat, od rýb cez vtáky až po cicavce, má endokanabinoid a vieme, že ľudia si nielen vyrábajú svoje vlastné kanabinoidy, ktoré interagujú s týmto konkrétnym systémom, ale produkujeme aj iné zlúčeniny, ktoré interagujú s ECS, napr. ktoré sú pozorované v mnohých rôznych rastlinách a potravinách, ďaleko za druhmi kanabisu.
Ako systém ľudského tela nie je ECS izolovanou štrukturálnou platformou, ako je nervový systém alebo kardiovaskulárny systém. Namiesto toho je ECS súborom receptorov široko distribuovaných v tele, ktoré sú aktivované prostredníctvom súboru ligandov, ktoré spoločne poznáme ako endokanabinoidy alebo endogénne kanabinoidy. Oba overené receptory sa nazývajú len CB1 a CB2, hoci existujú aj iné, ktoré boli navrhnuté. Kanály PPAR a TRP tiež sprostredkúvajú niektoré funkcie. Rovnako tak nájdete len dva dobre zdokumentované endokanabinoidy: anadamid a 2-arachidonoylglycerol alebo 2-AG.
Navyše, základom endokanabinoidného systému sú enzýmy, ktoré syntetizujú a rozkladajú endokanabinoidy. Predpokladá sa, že endokanabinoidy sa syntetizujú v základoch podľa potreby. Primárnymi enzýmami sú diacylglycerollipáza a N-acyl-fosfatidyletanolamín-fosfolipáza D, ktoré syntetizujú 2-AG a anandamid. Dva hlavné degradujúce enzýmy sú hydroláza amidu mastných kyselín alebo FAAH, ktorá rozkladá anandamid, a monoacylglycerollipáza alebo MAGL, ktorá rozkladá 2-AG. Regulácia týchto dvoch enzýmov môže zvýšiť alebo znížiť moduláciu ECS.
Aká je funkcia ECS?
ECS je hlavným homeostatickým regulačným systémom tela. Možno ho ľahko považovať za vnútorný adaptogénny systém tela, ktorý vždy pracuje na udržaní rovnováhy rôznych funkcií. Endokanabinoidy vo všeobecnosti fungujú ako neuromodulátory a ako také regulujú širokú škálu telesných procesov, od plodnosti až po bolesť. Niektoré z tých známejších funkcií z ECS sú nasledovné:
Nervový systém
Z centrálneho nervového systému alebo CNS bude všeobecná stimulácia CB1 receptorov inhibovať uvoľňovanie glutamátu a GABA. V CNS hrá ECS úlohu pri tvorbe pamäte a učení, podporuje neurogenézu v hipokampe a tiež reguluje excitabilitu neurónov. ECS tiež hrá úlohu v spôsobe, akým bude mozog reagovať na zranenie a zápal. Z miechy moduluje ECS signalizáciu bolesti a zvyšuje prirodzenú analgéziu. V periférnom nervovom systéme, v ktorom kontrolujú receptory CB2, pôsobí ECS primárne v sympatickom nervovom systéme na reguláciu funkcií črevného, močového a reprodukčného traktu.
Stres a nálada
ECS má viacero vplyvov na stresové reakcie a emocionálnu reguláciu, ako je iniciácia tejto telesnej reakcie na akútny stres a adaptácia v priebehu času na dlhodobejšie emócie, ako je strach a úzkosť. Zdravo fungujúci endokanabinoidný systém je rozhodujúci pre to, ako ľudia modulujú medzi uspokojivým stupňom vzrušenia v porovnaní s úrovňou, ktorá je nadmerná a nepríjemná. ECS tiež zohráva úlohu pri formovaní pamäte a možno najmä v spôsobe, akým si mozog vtláča spomienky zo stresu alebo zranenia. Pretože ECS moduluje uvoľňovanie dopamínu, noradrenalínu, serotonínu a kortizolu, môže tiež vo veľkej miere ovplyvniť emocionálnu reakciu a správanie.
Zažívacie ústrojenstvo
Tráviaci trakt je obsadený receptormi CB1 aj CB2, ktoré regulujú niekoľko dôležitých aspektov zdravia GI. Predpokladá sa, že ECS môže byť „chýbajúcim článkom“ pri opise prepojenia čreva, mozgu a imunity, ktoré hrá významnú úlohu vo funkčnom zdraví tráviaceho traktu. ECS je regulátorom imunity čriev, možno tým, že obmedzuje imunitný systém v ničení zdravej flóry a tiež prostredníctvom modulácie signalizácie cytokínov. ECS moduluje prirodzenú zápalovú odpoveď v tráviacom trakte, čo má dôležité dôsledky pre širokú škálu zdravotných problémov. Zdá sa, že motilita žalúdka a celková GI motilita je tiež čiastočne riadená ECS.
Chuť do jedla a metabolizmus
ECS, najmä receptory CB1, hrá úlohu pri chuti do jedla, metabolizme a regulácii telesného tuku. Stimulácia receptorov CB1 zvyšuje správanie pri hľadaní potravy, zlepšuje vnímanie vône a tiež reguluje energetickú rovnováhu. Zvieratá aj ľudia s nadváhou majú dysreguláciu ECS, ktorá môže viesť k tomu, že sa tento systém stane hyperaktívnym, čo prispieva k prejedaniu a zníženiu výdaja energie. Ukázalo sa, že cirkulujúce hladiny anandamidu a 2-AG sú zvýšené pri obezite, čo môže byť čiastočne spôsobené zníženou produkciou enzýmu degradujúceho FAAH.
Imunitné zdravie a zápalová reakcia
Bunky a orgány imunitného systému sú bohaté na endokanabinoidné receptory. Kanabinoidné receptory sú exprimované v týmuse, slezine, mandlích a kostnej dreni, ako aj na T- a B-lymfocytoch, makrofágoch, žírnych bunkách, neutrofiloch a prirodzených zabíjačských bunkách. ECS sa považuje za primárnu hnaciu silu rovnováhy imunitného systému a homeostázy. Hoci nie sú pochopené všetky funkcie ECS z imunitného systému, zdá sa, že ECS reguluje produkciu cytokínov a má tiež úlohu pri prevencii nadmernej aktivity imunitného systému. Zápal je prirodzenou súčasťou imunitnej odpovede a hrá veľmi normálnu úlohu pri akútnych poškodeniach tela vrátane zranení a chorôb; napriek tomu, keď nie je pod kontrolou, môže sa stať chronickým a prispievať ku kaskáde nepriaznivých zdravotných problémov, ako je chronická bolesť. Tým, že ECS udržiava imunitnú odpoveď pod kontrolou, pomáha udržiavať vyváženejšiu zápalovú odpoveď v tele.
Ďalšie oblasti zdravia regulované ECS:
Zdravie kostí
plodnosť
zdravie Skin
Zdravie artérií a dýchacích ciest
Spánok a cirkadiánny rytmus
Ako najlepšie podporiť zdravé ECS je otázka, na ktorú sa teraz mnohí výskumníci snažia odpovedať. Zostaňte naladení pre ďalšie informácie o tejto vznikajúcej téme.
Záverom možno povedať,Chronická bolesť je spojená so zmenami mozgu, vrátane redukcie šedej hmoty. Vyššie uvedený článok však ukázal, že chronická bolesť môže zmeniť celkovú štruktúru a funkciu mozgu. Hoci chronická bolesť môže viesť k týmto, okrem iných zdravotných problémov, správna liečba základných symptómov pacienta môže zvrátiť zmeny mozgu a regulovať šedú hmotu. Okrem toho sa čoraz viac výskumných štúdií objavuje za dôležitosťou endokanabinoidného systému a jeho funkcie pri kontrole, ako aj pri zvládaní chronickej bolesti a iných zdravotných problémov. Informácie, na ktoré odkazuje Národné centrum pre biotechnologické informácie (NCBI).�Rozsah našich informácií je obmedzený na chiroprax, ako aj na poranenia a stavy chrbtice. Ak chcete prediskutovať túto tému, neváhajte sa opýtať Dr. Jimeneza alebo nás kontaktujte na adrese�915-850-0900 .
Kurátorom je Dr. Alex Jimenez
Ďalšie témy: Bolesť chrbta
Bolesti chrbta je jednou z najčastejších príčin invalidity a vymeškaných dní v práci na celom svete. Bolesti chrbta sa v skutočnosti pripisujú ako druhý najčastejší dôvod návštevy lekára, prevyšujú ju len infekcie horných dýchacích ciest. Približne 80 percent populácie aspoň raz za život zažije nejaký typ bolesti chrbta. Chrbtica je zložitá štruktúra pozostávajúca z kostí, kĺbov, väzov a svalov, okrem iného z mäkkých tkanív. Z tohto dôvodu môžu zranenia a/alebo zhoršené stavy, ako napr herné disky, môže nakoniec viesť k príznakom bolesti chrbta. Športové zranenia alebo zranenia pri automobilových nehodách sú často najčastejšou príčinou bolesti chrbta, niekedy však môžu mať aj tie najjednoduchšie pohyby bolestivé následky. Našťastie alternatívne možnosti liečby, ako je chiropraktická starostlivosť, môžu pomôcť zmierniť bolesť chrbta pomocou úprav chrbtice a manuálnych manipulácií, čo v konečnom dôsledku zlepšuje úľavu od bolesti.
3.�Wrigley PJ, Gustin SM, Macey PM, Nash PG, Gandevia SC a kol. (2009)�Anatomické zmeny v ľudskej motorickej kôre a motorických dráhach po úplnom poranení hrudnej miechy.�Cereb Cortex�19: 224�232.�[PubMed]
4.�Máj A (2008)�Chronická bolesť môže zmeniť štruktúru mozgu.�Bolesť�137: 7�15.�[PubMed]
5.�Máj A (2009) Morfovanie voxelov: humbuk okolo štrukturálneho zobrazovania pacientov s bolesťou hlavy. Mozog.[PubMed]
6.�Apkarian AV, Baliki MN, Geha PY (2009)�Smerom k teórii chronickej bolesti.�Prog Neurobiol�87: 81�97.�[Článok bez PMC]�[PubMed]
7.�Apkarian AV, Sosa Y, Sonty S, Levy RM, Harden RN a kol. (2004)�Chronická bolesť chrbta je spojená so zníženou hustotou prefrontálnej a talamickej šedej hmoty.�J Neurosci�24: 10410�10415.�[PubMed]
8.�Rocca MA, Ceccarelli A, Falini A, Colombo B, Tortorella P a kol. (2006)�Zmeny sivej hmoty mozgu u pacientov s migrénou s T2-viditeľnými léziami: štúdia 3-T MRI.�Mŕtvica�37: 1765�1770.�[PubMed]
9.�Kuchinad A, Schweinhardt P, Seminowicz DA, Wood PB, Chizh BA a kol. (2007)�Zrýchlená strata sivej hmoty mozgu u pacientov s fibromyalgiou: predčasné starnutie mozgu?�J Neurosci�27: 4004 - 4007.[PubMed]
10.�Tracey I, Bushnell MC (2009)�Ako nás neuroimagingové štúdie vyzvali, aby sme prehodnotili: je chronická bolesť choroba?�J Bolesť�10: 1113�1120.�[PubMed]
11.�Franke K, Ziegler G, Kloppel S, Gaser C (2010)�Odhadovanie veku zdravých jedincov z T1-vážených MRI skenov pomocou metód jadra: skúmanie vplyvu rôznych parametrov.�Neuroimage�50: 883�892.�[PubMed]
12.�Draganski B, máj A (2008)�Štrukturálne zmeny vyvolané tréningom v mozgu dospelého človeka.�Behav Brain Res�192: 137�142.�[PubMed]
13.�Adkins DL, Boychuk J, Remple MS, Kleim JA (2006)�Motorický tréning vyvoláva špecifické vzorce plasticity v motorickej kôre a mieche.�J Appl Physiol�101: 1776�1782.�[PubMed]
14.�Duerden EG, Laverdure-Dupont D (2008)�Cvičenie robí kôru.�J Neurosci�28: 8655�8657.�[PubMed]
15.�Draganski B, Moser T, Lummel N, Ganssbauer S, Bogdahn U a kol. (2006)�Zníženie šedej hmoty talu po amputácii končatiny.�Neuroimage�31: 951�957.�[PubMed]
16.�Nikolajsen L, Brandsborg B, Lucht U, Jensen TS, Kehlet H (2006)�Chronická bolesť po totálnej endoprotéze bedrového kĺbu: celoštátna dotazníková štúdia.�Acta Anaesthesiol Scand�50: 495�500.�[PubMed]
17.�Rodriguez-Raecke R, Niemeier A, Ihle K, Ruether W, máj A (2009)�Pokles šedej hmoty mozgu pri chronickej bolesti je dôsledkom a nie príčinou bolesti.�J Neurosci�29: 13746�13750.�[PubMed]
18.�Beck AT, Ward CH, Mendelson M, Mock J, Erbaugh J (1961)�Súpis na meranie depresie.�Arch Gen Psychiatry�4: 561�571.�[PubMed]
19.�Franke G (2002) Príznakový kontrolný zoznam podľa LR Derogatis – Manuál. G�ttingen Beltz Test Verlag.
20.�Geissner E (1995) The Pain Perception Scale – diferencovaná a na zmeny citlivá škála na hodnotenie chronickej a akútnej bolesti. Rehabilitácia (Stuttg) 34: XXXV�XLIII.�[PubMed]
21.�Bullinger M, Kirchberger I (1998) SF-36 – Fragebogen zum Gesundheitszustand. Hand-anweisung. G�ttingen: Hogrefe.
22.�Ashburner J, Friston KJ (2000)�Morfometria založená na voxeloch.�Neuroimage�11: 805 - 821.[PubMed]
23.�Dobré CD, Johnsrude IS, Ashburner J, Henson RN, Friston KJ a spol. (2001)�Morfometrická štúdia starnutia založená na voxeloch v 465 normálnych mozgoch dospelých ľudí.�Neuroimage�14: 21�36.�[PubMed]
24.�Baliki MN, Chialvo DR, Geha PY, Levy RM, Harden RN a kol. (2006)�Chronická bolesť a emocionálny mozog: špecifická mozgová aktivita spojená so spontánnymi výkyvmi intenzity chronickej bolesti chrbta.�J Neurosci�26: 12165�12173.�[Článok bez PMC]�[PubMed]
25.�Lutz J, Jager L, de Quervain D, Krauseneck T, Padberg F a kol. (2008)�Abnormality bielej a šedej hmoty v mozgu pacientov s fibromyalgiou: štúdia difúzneho tenzora a volumetrického zobrazovania.�Arthritis Rheum�58: 3960�3969.�[PubMed]
26.�Wrigley PJ, Gustin SM, Macey PM, Nash PG, Gandevia SC a kol. (2008)�Anatomické zmeny v ľudskej motorickej kôre a motorických dráhach po úplnom poranení hrudnej miechy.�Cereb Cortex19: 224�232.�[PubMed]
27.�Schmidt-Wilcke T, Hierlmeier S, Leinisch E (2010) Zmenená regionálna morfológia mozgu u pacientov s chronickou bolesťou tváre. Bolesť hlavy.�[PubMed]
28.�Geha PY, Baliki MN, Harden RN, Bauer WR, Parrish TB a kol. (2008)�Mozog pri chronickej bolesti CRPS: abnormálne interakcie šedo-bielej hmoty v emocionálnych a autonómnych oblastiach.�Neurón�60: 570�581.�[Článok bez PMC]�[PubMed]
29.�Brazier J, Roberts J, Deverill M (2002)�Odhad miery zdravia na základe preferencie z SF-36.�J Health Econ�21: 271�292.�[PubMed]
30.�Draganski B, Gaser C, Busch V, Schuierer G, Bogdahn U a kol. (2004)�Neuroplasticita: zmeny šedej hmoty vyvolané tréningom.�príroda�427: 311�312.�[PubMed]
31.�Boyke J, Driemeyer J, Gaser C, Buchel C, máj A (2008)�Zmeny v štruktúre mozgu vyvolané tréningom u starších ľudí.�J Neurosci�28: 7031�7035.�[PubMed]
32.�Driemeyer J, Boyke J, Gaser C, Buchel C, máj A (2008)�Zmeny v sivej hmote vyvolané učením sa prehodnotili.�PLoS ONE�3: e2669.�[Článok bez PMC]�[PubMed]
33.�May A, Hajak G, Ganssbauer S, Steffens T, Langguth B a kol. (2007)�Štrukturálne zmeny mozgu po 5 dňoch intervencie: dynamické aspekty neuroplasticity.�Cereb Cortex�17: 205�210.�[PubMed]
34.�Teutsch S, Herken W, Bingel U, Schoell E, máj A (2008)�Zmeny sivej hmoty mozgu v dôsledku opakovanej bolestivej stimulácie.�Neuroimage�42: 845�849.�[PubMed]
35.�Flor H, Braun C, Elbert T, Birbaumer N (1997)�Rozsiahla reorganizácia primárnej somatosenzorickej kôry u pacientov s chronickou bolesťou chrbta.�Neurosci Lett�224: 5�8.�[PubMed]
36.�Flor H, Denke C, Schaefer M, Grusser S (2001)�Vplyv tréningu senzorickej diskriminácie na kortikálnu reorganizáciu a fantómovú bolesť končatín.�Lanceta�357: 1763�1764.�[PubMed]
37.�Swart CM, Stins JF, Beek PJ (2009)�Kortikálne zmeny pri syndróme komplexnej regionálnej bolesti (CRPS).�Eur J Pain�13: 902�907.�[PubMed]
38.�Maihofner C, Baron R, DeCol R, Binder A, Birklein F, a kol. (2007)�Motorický systém vykazuje adaptívne zmeny v syndróme komplexnej regionálnej bolesti.�Mozog�130: 2671�2687.�[PubMed]
39.�Fontaine D, Hamani C, Lozano A (2009)�Účinnosť a bezpečnosť stimulácie motorickej kôry pre chronickú neuropatickú bolesť: kritický prehľad literatúry.�J Neurosurg�110: 251�256.�[PubMed]
40.�Levy R, Deer TR, Henderson J (2010)�Intrakraniálna neurostimulácia na kontrolu bolesti: prehľad.�Lekársky lekár�13: 157�165.�[PubMed]
41.�Antal A, Brepohl N, Poreisz C, Boros K, Csifcsak G, a kol. (2008)�Transkraniálna stimulácia jednosmerným prúdom cez somatosenzorickú kôru znižuje experimentálne indukované vnímanie akútnej bolesti.�Clin J Pain24: 56�63.�[PubMed]
42.�Teepker M, Hotzel J, Timmesfeld N, Reis J, Mylius V a kol. (2010)�Nízkofrekvenčná rTMS vertexu v profylaktickej liečbe migrény.�Cephalalgia�30: 137�144.�[PubMed]
43.�O�Connell N, Wand B, Marston L, Spencer S, Desouza L (2010)�Neinvazívne techniky stimulácie mozgu pre chronickú bolesť. Správa Cochranovho systematického prehľadu a metaanalýzy.�Eur. J. Phys. Rehabil. Med�47: 309�326.�[PubMed]
44.�Tsao H, Galea MP, Hodges PW (2008)�Reorganizácia motorickej kôry je spojená s deficitom posturálnej kontroly pri opakovaných bolestiach krížov.�Mozog�131: 2161�2171.�[PubMed]
45.�Puri BK, Agour M, Gunatilake KD, Fernando KA, Gurusinghe AI a kol. (2010)�Zníženie šedej hmoty ľavej doplnkovej motorickej oblasti u dospelých žien trpiacich fibromyalgiou s výraznou únavou a bez afektívnej poruchy: pilotná kontrolovaná morfometrická štúdia 3-T magnetickou rezonanciou založená na voxeli.�J Int Med Res�38: 1468�1472.�[PubMed]
46.�Gwilym SE, Fillipini N, Douaud G, Carr AJ, Tracey I (2010) Talamická atrofia spojená s bolestivou osteoartritídou bedrového kĺbu je po artroplastike reverzibilná; longitudinálna morfometrická štúdia na báze voxelov. Arthritis Rheum.�[PubMed]
47.�Seminowicz DA, Wideman TH, Naso L, Hatami-Khoroushahi Z, Fallatah S, a kol. (2011)�Účinná liečba chronickej bolesti dolnej časti chrbta u ľudí zvráti abnormálnu anatómiu a funkciu mozgu.�J Neurosci31: 7540�7550.�[PubMed]
48.�May A, Gaser C (2006)�Morfometria založená na magnetickej rezonancii: okno do štrukturálnej plasticity mozgu.�Curr Opin Neurol�19: 407�411.�[PubMed]
49.�Schmidt-Wilcke T, Leinisch E, Straube A, Kampfe N, Draganski B a kol. (2005)�Pokles šedej hmoty u pacientov s chronickou bolesťou hlavy typu napätia.�Neurológia�65: 1483�1486.�[PubMed]
50.�Máj A (2009)�Morfovanie voxelov: humbuk okolo štrukturálneho zobrazovania pacientov s bolesťou hlavy.�Mozog 132 (Pt6): 1419�1425.�[PubMed]
Biochémia bolesti:�Všetky bolestivé syndrómy majú zápalový profil. Zápalový profil sa môže líšiť od osoby k osobe a môže sa líšiť aj u jednej osoby v rôznych časoch. Liečba bolestivých syndrómov spočíva v pochopení tohto zápalového profilu. Bolestivé syndrómy sa liečia medicínsky, chirurgicky alebo oboje. Cieľom je inhibovať/potlačiť produkciu zápalových mediátorov. A úspešný výsledok je ten, ktorý vedie k menšiemu zápalu a samozrejme menšej bolesti.
Biochémia bolesti
ciele:
Kto sú kľúčoví hráči
Aké sú biochemické mechanizmy?
Aké sú dôsledky?
Hodnotenie zápalu:
kľúčoví hráči
Prečo ma bolí rameno? Prehľad neuroanatomického a biochemického základu bolesti ramena
Abstrakt
Ak sa pacient spýta �prečo ma bolí rameno?� rozhovor sa rýchlo zvrtne na vedeckú teóriu a niekedy nepodložené dohady. Kliník si často uvedomuje limity vedeckého základu ich vysvetlenia, čo poukazuje na neúplnosť nášho chápania povahy bolesti ramena. Tento prehľad využíva systematický prístup, ktorý pomáha odpovedať na základné otázky týkajúce sa bolesti ramena, s cieľom poskytnúť pohľad na budúci výskum a nové metódy liečby bolesti ramena. Preskúmame úlohy (1) periférnych receptorov, (2) periférneho spracovania bolesti alebo „nocicepcie“, (3) miechy, (4) mozgu, (5) umiestnenia receptorov v ramene a (6 ) nervová anatómia ramena. Zvažujeme tiež, ako môžu tieto faktory prispieť k variabilite v klinickom obraze, diagnostike a liečbe bolesti ramena. Týmto spôsobom sa snažíme poskytnúť prehľad komponentov periférneho systému detekcie bolesti a centrálnych mechanizmov spracovania bolesti pri bolestiach ramena, ktoré interagujú a vytvárajú klinickú bolesť.
ÚVOD: VEĽMI STRUČNÁ HISTÓRIA VEDY BOLESTI NEZBYTNÁ PRE KLINIKOV
Povaha bolesti bola vo všeobecnosti v minulom storočí predmetom mnohých kontroverzií. V 17. storočí Descartova teória1 navrhovala, že intenzita bolesti priamo súvisí s množstvom súvisiaceho poranenia tkaniva a že bolesť sa spracováva jednou odlišnou cestou. Mnoho skorších teórií sa opieralo o túto takzvanú „dualistickú“ Descartovskú filozofiu, ktorá videla bolesť ako dôsledok stimulácie „špecifického“ periférneho receptora bolesti v mozgu. V 20. storočí nasledoval vedecký boj medzi dvoma protichodnými teóriami, konkrétne teóriou špecifickosti a teóriou vzorov. Descartovská „teória špecifickosti“ videla bolesť ako špecifickú samostatnú modalitu zmyslového vstupu s vlastným aparátom, zatiaľ čo „teória vzorcov“ sa domnievala, že bolesť je výsledkom intenzívnej stimulácie nešpecifických receptorov.2 V roku 1965 Wall a Melzack 3 teória bolesti poskytla dôkaz pre model, v ktorom bolo vnímanie bolesti modulované ako senzorickou spätnou väzbou, tak aj centrálnym nervovým systémom. Ďalší obrovský pokrok v teórii bolesti približne v rovnakom čase zaznamenal objav špecifického mechanizmu účinku opioidov.4 Následne nedávne pokroky v neurozobrazovaní a molekulárnej medicíne výrazne rozšírili naše celkové chápanie bolesti.
Ako to teda súvisí s bolesťou ramena?�Bolesť ramena je bežným klinickým problémoma dôkladné pochopenie spôsobu, akým telo spracováva bolesť, je nevyhnutné na čo najlepšiu diagnostiku a liečbu pacientovej bolesti. Pokroky v našich znalostiach spracovania bolesti sľubujú vysvetliť nesúlad medzi patológiou a vnímaním bolesti, môžu nám tiež pomôcť vysvetliť, prečo niektorí pacienti nereagujú na určité liečby.
ZÁKLADNÉ STAVEBNÉ BLOKY BOLESTI
Periférne senzorické receptory: mechanoreceptor a „nociceptor“.
V ľudskom muskuloskeletálnom systéme je prítomných množstvo typov periférnych senzorických receptorov. 5 Môžu byť klasifikované na základe ich funkcie (ako mechanoreceptory, termoreceptory alebo nociceptory) alebo morfológie (voľné nervové zakončenia alebo rôzne typy zapuzdrených receptorov).5 Rôzne typy receptorov sa potom môžu ďalej klasifikovať na základe prítomnosť určitých chemických markerov. Napríklad existujú významné prekrytia medzi rôznymi funkčnými triedami receptorov
Spracovanie periférnej bolesti: �Nocicepcia�
Poškodenie tkaniva zahŕňa rôzne zápalové mediátory uvoľňované poškodenými bunkami vrátane bradykinínu, histamínu, 5-hydroxytryptamínu, ATP, oxidu dusnatého a určitých iónov (K+ a H+). Aktivácia dráhy kyseliny arachidónovej vedie k produkcii prostaglandínov, tromboxánov a leukotriénov. Cytokíny, vrátane interleukínov a tumor nekrotizujúceho faktora ?, a neurotrofíny, ako je nervový rastový faktor (NGF), sa tiež uvoľňujú a sú úzko zapojené do uľahčenia zápalu.15 Ďalšie látky, ako sú excitačné aminokyseliny (glutamát) a opioidy ( endotelín-1) sa tiež podieľajú na akútnej zápalovej odpovedi.16 17 Niektoré z týchto činidiel môžu priamo aktivovať nociceptory, zatiaľ čo iné spôsobujú nábor iných buniek, ktoré potom uvoľňujú ďalšie uľahčujúce činidlá.18 Tento lokálny proces vedie k zvýšenej citlivosti nociceptívnych neurónov na ich normálny vstup a/alebo nábor odpovede na normálne podprahové vstupy sa nazýva „periférna senzibilizácia“.�Obrázok 1 sumarizuje niektoré zo zahrnutých kľúčových mechanizmov.
NGF a receptor katiónového kanála s prechodným potenciálom receptora V člen 1 (TRPV1) majú symbiotický vzťah, pokiaľ ide o zápal a senzibilizáciu nociceptora. Cytokíny produkované v zapálenom tkanive vedú k zvýšeniu produkcie NGF.19 NGF stimuluje uvoľňovanie histamínu a serotonínu (5-HT3) žírnymi bunkami a tiež senzibilizuje nociceptory, čo môže zmeniť vlastnosti A? vlákna tak, že väčšia časť sa stáva nociceptívnou. Receptor TRPV1 je prítomný v subpopulácii primárnych aferentných vlákien a je aktivovaný kapsaicínom, teplom a protónmi. Receptor TRPV1 je syntetizovaný v bunkovom tele aferentného vlákna a je transportovaný do periférnych aj centrálnych zakončení, kde prispieva k senzitivite nociceptívnych aferentov. Zápal vedie k periférnej produkcii NGF, ktorý sa potom naviaže na receptor tyrozínkinázy typu 1 na zakončeniach nociceptora, NGF je potom transportovaný do bunkového tela, kde vedie k upregulácii transkripcie TRPV1 a následne k zvýšeniu citlivosti nociceptora.19 20 NGF a iné zápalové mediátory tiež senzibilizujú TRPV1 prostredníctvom rôznorodého radu sekundárnych mediátorových dráh. Mnoho ďalších receptorov vrátane cholinergných receptorov, receptorov kyseliny a-aminomaslovej (GABA) a somatostatínových receptorov sa tiež považuje za zapojené do periférnej citlivosti nociceptorov.
Veľký počet zápalových mediátorov sa špecificky podieľa na bolestiach ramena a ochorení rotátorovej manžety.21�25 Zatiaľ čo niektoré chemické mediátory priamo aktivujú nociceptory, väčšina vedie skôr k zmenám v samotnom senzorickom neuróne, než by ho priamo aktivovala. Tieto zmeny môžu byť závislé od skorej post-translačnej alebo oneskorenej transkripcie. Príkladmi prvých sú zmeny v receptore TRPV1 alebo v napäťovo riadených iónových kanáloch, ktoré sú výsledkom fosforylácie membránovo viazaných proteínov. Príklady posledne menovaných zahŕňajú zvýšenie produkcie kanálov TRV1 indukované NGF a aktiváciu intracelulárnych transkripčných faktorov indukovanú vápnikom.
Molekulárne mechanizmy nocicepcie
Pocit bolesti nás upozorňuje na skutočné alebo hroziace zranenie a spúšťa vhodné ochranné reakcie. Bohužiaľ, bolesť často prežije svoju užitočnosť ako varovný systém a namiesto toho sa stáva chronickou a oslabujúcou. Tento prechod do chronickej fázy zahŕňa zmeny v mieche a mozgu, ale existuje aj pozoruhodná modulácia, kde sa iniciujú správy o bolesti – na úrovni primárneho senzorického neurónu. Úsilie určiť, ako tieto neuróny detegujú podnety spôsobujúce bolesť tepelnej, mechanickej alebo chemickej povahy, odhalili nové signalizačné mechanizmy a priviedli nás bližšie k pochopeniu molekulárnych dejov, ktoré uľahčujú prechody od akútnej k pretrvávajúcej bolesti.
Neurochémia nociceptorov
Glutamát je prevládajúci excitačný neurotransmiter vo všetkých nociceptoroch. Histochemické štúdie dospelých DRG však odhaľujú dve široké triedy nemyelinizovaných vlákien C.
Chemické prevodníky na zhoršenie bolesti
Ako je opísané vyššie, zranenie zvyšuje našu bolesť tým, že zvyšuje citlivosť nociceptorov na tepelné aj mechanické stimuly. Tento jav je čiastočne výsledkom produkcie a uvoľňovania chemických mediátorov z primárneho senzorického zakončenia az neneurálnych buniek (napríklad fibroblastov, žírnych buniek, neutrofilov a krvných doštičiek) v prostredí36 (obr. 3). Niektoré zložky zápalovej polievky (napríklad protóny, ATP, serotonín alebo lipidy) môžu meniť neurónovú excitabilitu priamo interakciou s iónovými kanálmi na povrchu nociceptora, zatiaľ čo iné (napríklad bradykinín a NGF) sa viažu na metabotropné receptory a sprostredkúvajú svoje účinky prostredníctvom signálnych kaskád druhého posla11. V pochopení biochemického základu takýchto modulačných mechanizmov sa dosiahol značný pokrok.
Extracelulárne protóny a acidóza tkanív
Lokálna acidóza tkaniva je charakteristickou fyziologickou odpoveďou na poranenie a stupeň súvisiacej bolesti alebo nepohodlia dobre koreluje s veľkosťou acidifikácie37. Aplikácia kyseliny (pH 5) na kožu vytvára trvalé výboje v tretine alebo viacerých polymodálnych nociceptoroch, ktoré inervujú receptívne pole 20.
Bunkové a molekulárne mechanizmy bolesti
abstraktné
Nervový systém detekuje a interpretuje širokú škálu tepelných a mechanických stimulov, ako aj environmentálnych a endogénnych chemických dráždidiel. Keď sú tieto stimuly intenzívne, generujú akútnu bolesť a pri pretrvávajúcom poškodení vykazujú zložky periférneho aj centrálneho nervového systému dráhy prenosu bolesti obrovskú plasticitu, zosilňujú signály bolesti a spôsobujú precitlivenosť. Keď plasticita uľahčuje ochranné reflexy, môže to byť prospešné, ale keď zmeny pretrvávajú, môže to viesť k chronickej bolesti. Genetické, elektrofyziologické a farmakologické štúdie objasňujú molekulárne mechanizmy, ktoré sú základom detekcie, kódovania a modulácie škodlivých stimulov, ktoré spôsobujú bolesť.
Úvod: Akútna verzus pretrvávajúca bolesť
Obrázok 5. Miechová (centrálna) senzibilizácia
Glutamát/NMDA receptorom sprostredkovaná senzibilizácia.�Po intenzívnej stimulácii alebo pretrvávajúcom poranení sa aktivovali C a A? nociceptory uvoľňujú rôzne neurotransmitery vrátane dlutamátu, substancie P, peptidu súvisiaceho s génom kalcitonínu (CGRP) a ATP na výstupné neuróny v lamine I povrchového dorzálneho rohu (červená). V dôsledku toho môžu normálne tiché glutamátové receptory NMDA umiestnené v postsynaptickom neuróne teraz signalizovať, zvyšovať intracelulárny vápnik a aktivovať množstvo signálnych dráh závislých od vápnika a druhých poslov vrátane mitogénom aktivovanej proteínkinázy (MAPK), proteínkinázy C (PKC) proteínkináza A (PKA) a Src. Táto kaskáda udalostí zvýši excitabilitu výstupného neurónu a uľahčí prenos správ o bolesti do mozgu.
Dezinhibícia.�Za normálnych okolností inhibičné interneuróny (modré) nepretržite uvoľňujú GABA a/alebo glycín (Gly), aby sa znížila excitabilita výstupných neurónov laminy I a modulovali sa prenos bolesti (inhibičný tón). V prípade poranenia sa však táto inhibícia môže stratiť, čo vedie k hyperalgézii. Okrem toho môže dezinhibícia umožniť nenociceptívny myelinizovaný Ap. primárne aferenty na zapojenie obvodu prenosu bolesti tak, že normálne neškodné stimuly sú teraz vnímané ako bolestivé. K tomu dochádza čiastočne prostredníctvom dezinhibície excitačnej PKC? exprimujúce interneuróny vo vnútornej vrstve II.
Aktivácia mikroglií.�Poranenie periférnych nervov podporuje uvoľňovanie ATP a chemokínu fraktalkínu, ktorý stimuluje mikrogliálne bunky. Najmä aktivácia purinergných, CX3CR1 a Toll-like receptorov na mikrogliách (fialová) má za následok uvoľnenie neurotrofického faktora odvodeného z mozgu (BDNF), ktorý prostredníctvom aktivácie TrkB receptorov exprimovaných výstupnými neurónmi lamina I podporuje zvýšenú excitabilitu a zvýšená bolesť v reakcii na škodlivú aj neškodnú stimuláciu (to znamená hyperalgézia a alodýnia). Aktivované mikroglie tiež uvoľňujú množstvo cytokínov, ako je tumor nekrotizujúci faktor ? (TNFa), interleukín-1? a 6 (IL-1p, IL-6) a ďalšie faktory, ktoré prispievajú k centrálnej senzibilizácii.
Chemické prostredie zápalu
Periférna senzibilizácia je častejšie výsledkom zmien spojených so zápalom v chemickom prostredí nervového vlákna (McMahon et al., 2008). Poškodenie tkaniva je teda často sprevádzané akumuláciou endogénnych faktorov uvoľnených z aktivovaných nociceptorov alebo neneurálnych buniek, ktoré sa nachádzajú v poranenej oblasti alebo do nej infiltrujú (vrátane žírnych buniek, bazofilov, krvných doštičiek, makrofágov, neutrofilov, endotelových buniek, keratinocytov a fibroblasty). kolektívne. tieto faktory, označované ako „zápalová polievka“, predstavujú širokú škálu signálnych molekúl, vrátane neurotransmiterov, peptidov (látka P, CGRP, bradykinín), eikozinoidov a príbuzných lipidov (prostaglandíny, tromboxány, leukotriény, endokanabinoidy), neurotrofínov, cytokínov a chemokíny, ako aj extracelulárne proteázy a protóny. Je pozoruhodné, že nociceptory exprimujú jeden alebo viac bunkových povrchových receptorov schopných rozpoznať a reagovať na každé z týchto prozápalových alebo proalgetických činidiel (obrázok 4). Takéto interakcie zvyšujú excitabilitu nervového vlákna, čím zvyšujú jeho citlivosť na teplotu alebo dotyk.
Nepochybne najbežnejší prístup k zníženiu zápalovej bolesti zahŕňa inhibíciu syntézy alebo akumulácie zložiek zápalovej polievky. Najlepším príkladom toho sú nesteroidné protizápalové lieky, ako je aspirín alebo ibuprofén, ktoré znižujú zápalovú bolesť a hyperalgéziu inhibíciou cyklooxygenáz (Cox-1 a Cox-2), ktoré sa podieľajú na syntéze prostaglandínov. Druhým prístupom je blokovanie pôsobenia zápalových činidiel na nociceptore. Tu zdôrazňujeme príklady, ktoré poskytujú nový pohľad na bunkové mechanizmy periférnej senzibilizácie alebo ktoré tvoria základ nových terapeutických stratégií na liečbu zápalovej bolesti.
NGF je možno najlepšie známy pre svoju úlohu neurotrofického faktora potrebného na prežitie a vývoj senzorických neurónov počas embryogenézy, ale u dospelých sa NGF produkuje aj pri poškodení tkaniva a tvorí dôležitú zložku zápalovej polievky (Ritner et al., 2009). Spomedzi svojich mnohých bunkových cieľov pôsobí NGF priamo na nociceptory peptidergických vlákien C, ktoré exprimujú vysoko afinitnú receptorovú tyrozínkinázu NGF, TrkA, ako aj nízkoafinitný neurotrofínový receptor p75 (Chao, 2003; Snider a McMahon, 1998). NGF spôsobuje hlbokú precitlivenosť na teplo a mechanické stimuly prostredníctvom dvoch časovo odlišných mechanizmov. Najprv interakcia NGF-TrkA aktivuje downstream signálne dráhy, vrátane fosfolipázy C (PLC), mitogénom aktivovanej proteínkinázy (MAPK) a fosfoinozitid-3-kinázy (PI3K). To vedie k funkčnej potenciácii cieľových proteínov na periférnom konci nociceptoru, najmä TRPV1, čo vedie k rýchlej zmene bunkovej a behaviorálnej citlivosti na teplo (Chuang et al., 2001).
Bez ohľadu na ich pro-nociceptívne mechanizmy sa interferencia s neurotrofínovou alebo cytokínovou signalizáciou stala hlavnou stratégiou kontroly zápalového ochorenia alebo výslednej bolesti. Hlavný prístup zahŕňa blokovanie NGF alebo TNF-? pôsobenie neutralizačnej protilátky. V prípade TNF-a to bolo pozoruhodne účinné pri liečbe mnohých autoimunitných ochorení, vrátane reumatoidnej artritídy, čo viedlo k dramatickému zníženiu deštrukcie tkaniva a sprievodnej hyperalgézie (Atzeni et al., 2005). Pretože hlavné účinky NGF na dospelý nociceptor sa vyskytujú na pozadí zápalu, výhodou tohto prístupu je, že hyperalgézia sa zníži bez ovplyvnenia normálne vnímanie bolesti. Anti-NGF protilátky sú v súčasnosti skutočne v klinických skúškach na liečbu syndrómov zápalovej bolesti (Hefti et al., 2006).
Akútna bolesť je signalizovaná uvoľňovaním glutamátu z centrálnych zakončení nociceptorov, čím sa vytvárajú excitačné postsynaptické prúdy (EPSC) v neurónoch dorzálnych rohov druhého poriadku. K tomu dochádza predovšetkým prostredníctvom aktivácie postsynaptických AMPA a kainátových podtypov ionotropných glutamátových receptorov. Sumácia podprahových EPSC v postsynaptickom neuróne nakoniec povedie k spusteniu akčného potenciálu a prenosu správy o bolesti na neuróny vyššieho rádu.
Iné štúdie naznačujú, že zmeny v samotnom projekčnom neuróne prispievajú k inhibičnému procesu. Napríklad poškodenie periférnych nervov výrazne znižuje K+- Cl- kotransportér KCC2, ktorý je nevyhnutný na udržanie normálnych gradientov K+ a Cl- cez plazmatickú membránu (Coull et al., 2003). Downregulácia KCC2, ktorý je exprimovaný v projekčných neurónoch lamina I, vedie k posunu v gradiente Cl, takže aktivácia receptorov GABA-A depolarizuje, a nie hyperpolarizuje projekčné neuróny lamina I. To by zase zvýšilo excitabilitu a zvýšilo prenos bolesti. Farmakologická blokáda alebo downregulácia KCC2 sprostredkovaná siRNA u potkanov skutočne indukuje mechanickú alodýniu.
Zdieľať e-knihu
Zdroje:
Prečo ma bolí rameno? Prehľad neuroanatomického a biochemického základu bolesti ramena
Benjamin John Floyd Dean, Stephen Edward Gwilym, Andrew Jonathan Carr
Bunkové a molekulárne mechanizmy bolesti
Allan I. Basbaum1, Diana M. Bautista2, Gre?gory Scherrer1 a David Julius3
1Katedra anatómie, Kalifornská univerzita, San Francisco 94158
2Katedra molekulárnej a bunkovej biológie, Kalifornská univerzita, Berkeley CA 94720 3Katedra fyziológie, Kalifornská univerzita, San Francisco 94158
Molekulárne mechanizmy nocicepcie
David Julius* a Allan I. Basbaum�
*Katedra bunkovej a molekulárnej farmakológie a �Katedry anatómie a fyziológie a WM Keck Foundation Center for Integrative Neuroscience, University of California San Francisco, San Francisco, California 94143, USA (e-mail: julius@socrates.ucsf.edu)
Neurogénny zápal, alebo NI, je fyziologický proces, pri ktorom sa mediátory uvoľňujú priamo z kožných nervov, aby sa začala zápalová odpoveď. To vedie k vytvoreniu lokálnych zápalových reakcií vrátane erytému, opuchu, zvýšenia teploty, citlivosti a bolesti. Za uvoľňovanie týchto zápalových mediátorov sú do značnej miery zodpovedné jemné nemyelinizované aferentné somatické C-vlákna, ktoré reagujú na nízku intenzitu mechanickej a chemickej stimulácie.
Keď sú tieto nervové dráhy v kožných nervoch stimulované, uvoľňujú energetické neuropeptidy alebo látku P a peptid súvisiaci s génom kalcitonínu (CGRP) rýchlo do mikroprostredia, čím spúšťajú sériu zápalových reakcií. Existuje významný rozdiel v imunogénnom zápale, čo je úplne prvá ochranná a reparačná reakcia imunitného systému, keď patogén vstúpi do tela, zatiaľ čo neurogénny zápal zahŕňa priame spojenie medzi nervovým systémom a zápalovými reakciami. Aj keď neurogénny zápal a imunologický zápal môžu existovať súčasne, nie sú tieto dva klinicky rozlíšiteľné. Účelom nižšie uvedeného článku je diskutovať o mechanizme neurogénneho zápalu a úlohe periférneho nervového systému v obrane hostiteľa a imunopatológii.
Neurogénny zápal – Úloha periférneho nervového systému v obrane hostiteľa a v imunopatológii
abstraktné
Periférny nervový a imunitný systém sa tradične považuje za systém slúžiaci oddeleným funkciám. Táto línia je však čoraz viac rozmazaná novými pohľadmi na neurogénny zápal. Nociceptorové neuróny majú mnoho rovnakých molekulárnych ciest rozpoznávania nebezpečenstva ako imunitné bunky a v reakcii na nebezpečenstvo komunikuje periférny nervový systém priamo s imunitným systémom a vytvára integrovaný ochranný mechanizmus. Hustá inervačná sieť senzorických a autonómnych vlákien v periférnych tkanivách a vysoká rýchlosť nervovej transdukcie umožňuje rýchlu lokálnu a systémovú neurogénnu moduláciu imunity. Zdá sa, že periférne neuróny zohrávajú významnú úlohu aj pri imunitnej dysfunkcii pri autoimunitných a alergických ochoreniach. Preto pochopenie koordinovanej interakcie periférnych neurónov s imunitnými bunkami môže pokročiť v terapeutických prístupoch na zvýšenie obrany hostiteľa a potlačenie imunopatológie.
úvod
Pred dvetisíc rokmi Celsus definoval zápal ako zápal zahŕňajúci štyri hlavné znaky: Dolor (bolesť), Calor (teplo), Rubor (začervenanie) a Tumor (opuch), čo je pozorovanie naznačujúce, že aktivácia nervového systému bola uznaná ako neoddeliteľná súčasť zápal. Na bolesť sa však odvtedy myslelo hlavne ako na symptóm, a nie na účastníka vytvárania zápalu. V tejto perspektíve ukazujeme, že periférny nervový systém zohráva priamu a aktívnu úlohu pri modulácii vrodených a adaptívnych imunity, takže imunitné a nervové systémy môžu mať spoločnú integrovanú ochrannú funkciu v hostiteľskej obrane a reakcii na poškodenie tkaniva, zložitého interakcie, ktorá môže tiež viesť k patológii alergických a autoimunitných ochorení.
Prežitie organizmov kriticky závisí od schopnosti postaviť sa na obranu proti potenciálnemu poškodeniu spôsobenému poškodením tkaniva a infekciou. Obrana hostiteľa zahŕňa vyhýbavé správanie na odstránenie kontaktu s nebezpečným (škodlivým) prostredím (neurálna funkcia) a aktívnu neutralizáciu patogénov (imunitná funkcia). Tradične sa úloha imunitného systému v boji proti infekčným agens a pri oprave poranenia tkaniva považovala za celkom odlišnú od úlohy nervového systému, ktorý premieňa škodlivé environmentálne a vnútorné signály na elektrickú aktivitu, ktorá vytvára pocity a reflexy (obr. 1). Navrhujeme, aby tieto dva systémy boli vlastne súčasťami jednotného obranného mechanizmu. Somatosenzorický nervový systém je ideálne umiestnený na detekciu nebezpečenstva. Po prvé, všetky tkanivá, ktoré sú vysoko exponované vonkajšiemu prostrediu, ako sú epiteliálne povrchy kože, pľúc, močového a tráviaceho traktu, sú husto inervované nociceptormi, zmyslovými vláknami produkujúcimi vysoký prah bolesti. Po druhé, prenos škodlivých vonkajších stimulov je takmer okamžitý, rádovo rýchlejší ako mobilizácia vrodeného imunitného systému, a preto môže byť „prvou odozvou“ pri obrane hostiteľa.
Obrázok 1: Škodlivé podnety, mikrobiálne a zápalové rozpoznávacie dráhy spúšťajú aktiváciu periférneho nervového systému. Senzorické neuróny majú niekoľko prostriedkov na zisťovanie prítomnosti škodlivých/škodlivých stimulov. 1) Nebezpečné signálne receptory vrátane TRP kanálov, P2X kanálov a receptory molekulového vzoru spojeného s nebezpečenstvom (DAMP) rozpoznávajú exogénne signály z prostredia (napr. teplo, kyslosť, chemikálie) alebo endogénne signály nebezpečenstva uvoľnené počas traumy/poškodenia tkaniva (napr. ATP, kyselina močová, hydroxynonaly). 2) Receptory na rozpoznávanie vzorov (PRR), ako sú Toll-like receptory (TLR) a Nod-like receptory (NLR), rozpoznávajú patogénne molekulové vzory (PAMP), ktoré sa vylučujú inváziou baktérií alebo vírusov počas infekcie. 3) Cytokínové receptory rozpoznávajú faktory vylučované imunitnými bunkami (napr. IL-1beta, TNF-alfa, NGF), ktoré aktivujú mapové kinázy a iné signalizačné mechanizmy na zvýšenie excitability membrány.
Okrem ortodromických vstupov do miechy a mozgu z periférie sa akčné potenciály v nociceptorových neurónoch môžu prenášať aj antidromicky v bodoch vetvenia späť na perifériu, axónový reflex. Tieto spolu s trvalými lokálnymi depolarizáciami vedú k rýchlemu a lokálnemu uvoľneniu nervových mediátorov z periférnych axónov aj terminálov (obr. 2). indukuje vazodilatáciu kože, čo viedlo ku konceptu „neurogénneho zápalu“ nezávislého od zápalu produkovaného imunitným systémom (obr. 1).
Obrázok 2: Neurónové faktory uvoľňované z nociceptorových senzorických neurónov priamo riadia chemotaxiu leukocytov, vaskulárnu hemodynamiku a imunitnú odpoveď. Keď škodlivé stimuly aktivujú aferentné signály v senzorických nervoch, vytvárajú sa antidromické axónové reflexy, ktoré indukujú uvoľňovanie neuropeptidov na periférnych zakončeniach neurónov. Tieto molekulárne mediátory majú niekoľko zápalových účinkov: 1) Chemotaxia a aktivácia neutrofilov, makrofágov a lymfocytov v mieste poškodenia a degranulácia žírnych buniek. 2) Signalizácia vaskulárnym endotelovým bunkám na zvýšenie prietoku krvi, vaskulárne presakovanie a edém. To tiež umožňuje ľahší nábor zápalových leukocytov. 3) Priming dendritických buniek na riadenie následnej diferenciácie pomocných T buniek na podtypy Th2 alebo Th17.
Obrázok 3: Časová os pokrokov v chápaní neurogénnych aspektov zápalu od Celsa po súčasnosť.
Neurogénny zápal je sprostredkovaný uvoľňovaním neuropeptidov, peptidu súvisiaceho s génom kalcitonínu (CGRP) a substancie P (SP) z nociceptorov, ktoré pôsobia priamo na vaskulárne endotelové bunky a bunky hladkého svalstva 2�5. CGRP vyvoláva vazodilatačné účinky 2, 3, zatiaľ čo SP zvyšuje kapilárnu permeabilitu, čo vedie k extravazácii plazmy a edému 4, 5, čo prispieva k ruboru, kalorácii a nádoru Celsus. Nociceptory však uvoľňujú mnoho ďalších neuropeptidov (online databáza: www.neuropeptides.nl/), vrátane adrenomedulínu, neurokinínov A a B, vazoaktívneho črevného peptidu (VIP), neuropeptidu (NPY) a peptidu uvoľňujúceho gastrín (GRP), ako aj iných molekulárnych mediátorov, ako je glutamát, oxid dusnatý (NO) a cytokíny, ako je eotaxín 6.
Teraz si uvedomujeme, že mediátory uvoľnené zo senzorických neurónov na periférii pôsobia nielen na vaskulatúru, ale tiež priamo priťahujú a aktivujú vrodené imunitné bunky (žírne bunky, dendritické bunky) a adaptívne imunitné bunky (T lymfocyty) 7�12. Pri akútnom poškodení tkaniva sa domnievame, že neurogénny zápal je ochranný, uľahčuje fyziologické hojenie rán a imunitnú obranu proti patogénom aktiváciou a náborom imunitných buniek. Takáto neuroimunitná komunikácia však pravdepodobne tiež hrá hlavnú úlohu v patofyziológii alergických a autoimunitných ochorení zosilnením patologických alebo maladaptívnych imunitných reakcií. Napríklad na zvieracích modeloch reumatoidnej artritídy Levine a kolegovia ukázali, že denervácia kĺbu vedie k výraznému oslabeniu zápalu, ktorý je závislý od nervovej expresie substancie P 13, 14. V nedávnych štúdiách alergických zápalov dýchacích ciest, kolitídy a psoriáza, primárne senzorické neuróny hrajú ústrednú úlohu pri iniciovaní a zvyšovaní aktivácie vrodenej a adaptívnej imunity 15�17.
Navrhujeme preto, aby periférny nervový systém zohrával nielen pasívnu úlohu pri obrane hostiteľa (detekcia škodlivých stimulov a iniciovanie vyhýbavého správania), ale aj aktívnu úlohu v zhode s imunitným systémom pri modulácii odpovedí na škodlivé látky a boji proti nim. stimuly, úlohu, ktorá môže byť narušená, aby prispela k chorobe.
Spoločné cesty rozpoznávania nebezpečenstva v periférnych nervových a vrodených imunitných systémoch
Periférne senzorické neuróny sú prispôsobené na rozpoznávanie nebezpečenstva pre organizmus na základe ich citlivosti na intenzívne mechanické, tepelné a dráždivé chemické podnety (obr. 1). Iónové kanály s prechodným receptorovým potenciálom (TRP) sú najviac študovanými molekulárnymi mediátormi nocicepcie, ktoré vedú neselektívny vstup katiónov po aktivácii rôznymi škodlivými stimulmi. TRPV1 je aktivovaný vysokými teplotami, nízkym pH a kapsaicínom, vallinoidnou dráždivou zložkou chilli papričiek 18. TRPA1 sprostredkováva detekciu reaktívnych chemikálií vrátane environmentálnych dráždivých látok, ako je slzný plyn a priemyselné izotiokyanáty 19, ale čo je dôležitejšie, aktivuje sa aj počas tkaniva poranenie endogénnymi molekulárnymi signálmi vrátane 4-hydroxynonenalu a prostaglandínov 20, 21.
Je zaujímavé, že senzorické neuróny zdieľajú mnohé z rovnakých patogénnych a nebezpečných molekulárnych receptorových dráh ako bunky vrodenej imunity, čo im umožňuje tiež detekovať patogény (obr. 1). V imunitnom systéme sú mikrobiálne patogény detegované zárodočnými kódovanými receptormi na rozpoznávanie vzorov (PRR), ktoré rozpoznávajú široko konzervované molekulárne vzory spojené s exogénnym patogénom (PAMP). Prvé PRR, ktoré boli identifikované, boli členovia rodiny toll-like receptorov (TLR), ktoré sa viažu na kvasinky, komponenty bunkovej steny odvodené od baktérií a vírusovú RNA 22. Po aktivácii PRR sa aktivujú downstream signálne dráhy, ktoré indukujú produkciu a aktiváciu cytokínov adaptívnej imunity. Okrem TLR sú vrodené imunitné bunky aktivované počas poškodenia tkaniva endogénnymi signálmi nebezpečenstva, ktoré sú tiež známe ako molekulárne vzorce spojené s poškodením (DAMP) alebo alarmíny 23, 24. Tieto nebezpečné signály zahŕňajú HMGB1, kyselinu močovú a uvoľnené proteíny tepelného šoku. odumieraním buniek počas nekrózy, aktiváciou imunitných buniek počas neinfekčných zápalových reakcií.
PRR vrátane TLR 3, 4, 7 a 9 sú exprimované nociceptorovými neurónmi a stimulácia TLR ligandmi vedie k indukcii vnútorných prúdov a senzibilizácii nociceptorov na iné bolestivé stimuly 25�27. Okrem toho, aktivácia senzorických neurónov ligandom TLR7 imikvimodom vedie k aktivácii senzorickej dráhy špecifickej pre svrbenie 25. Tieto výsledky naznačujú, že bolesť a svrbenie spojené s infekciou môžu byť čiastočne spôsobené priamou aktiváciou neurónov faktormi odvodenými od patogénu, ktoré zase aktivovať imunitné bunky periférnym uvoľňovaním neurónových signálnych molekúl.
Hlavným DAMP/alarmínom uvoľneným počas bunkového poškodenia je ATP, ktorý je rozpoznávaný purinergnými receptormi tak na nociceptorových neurónoch, ako aj na imunitných bunkách 28�30. Purinergné receptory sa skladajú z dvoch rodín: receptory P2X, katiónové kanály riadené ligandom, a receptory P2Y, receptory spojené s G-proteínom. V nociceptorových neurónoch dochádza k rozpoznávaniu ATP prostredníctvom P2X3, čo vedie k rýchlej denzitizácii katiónových prúdov a bolesti 28, 30 (obr. 1), zatiaľ čo receptory P2Y prispievajú k aktivácii nociceptora senzibilizáciou TRP a napäťovo riadených sodíkových kanálov. V makrofágoch vedie väzba ATP k receptorom P2X7 k hyperpolarizácii a následnej aktivácii zápalu, molekulárneho komplexu dôležitého pri vytváraní IL-1beta a IL-18 29. Preto je ATP silným nebezpečným signálom, ktorý aktivuje periférne neuróny aj vrodené imunitu počas poranenia a niektoré dôkazy dokonca naznačujú, že neuróny exprimujú časti zápalového molekulárneho aparátu 31.
Odvrátenou stranou nebezpečných signálov v nociceptoroch je úloha TRP kanálov pri aktivácii imunitných buniek. TRPV2, homológ TRPV1 aktivovaný škodlivým teplom, je exprimovaný vo vysokých hladinách v bunkách vrodenej imunity. 32. Genetická ablácia TRPV2 viedla k defektom vo fagocytóze makrofágov a odstráneniu bakteriálnych infekcií. ich degranulácia 32. Zostáva určiť, či endogénne signály nebezpečenstva aktivujú imunitné bunky podobným spôsobom ako nociceptory.
Kľúčovým prostriedkom komunikácie medzi imunitnými bunkami a nociceptorovými neurónmi sú cytokíny. Po aktivácii cytokínových receptorov sa v senzorických neurónoch aktivujú dráhy prenosu signálu, čo vedie k downstream fosforylácii membránových proteínov vrátane TRP a napäťovo riadených kanálov (obr. 1). Výsledná senzibilizácia nociceptorov znamená, že normálne neškodné mechanické a tepelné stimuly môžu teraz nociceptory aktivovať. Interleukín 1 beta a TNF-alfa sú dva dôležité cytokíny uvoľňované bunkami vrodenej imunity počas zápalu. IL-1beta a TNF-alfa sú priamo snímané nociceptormi, ktoré exprimujú príbuzné receptory, indukujú aktiváciu mapových kináz p38, čo vedie k zvýšenej excitabilite membrány 34�36. Nervový rastový faktor (NGF) a prostaglandín E(2) sú tiež hlavné zápalové mediátory uvoľňované z imunitných buniek, ktoré pôsobia priamo na periférne senzorické neuróny a spôsobujú senzibilizáciu. Dôležitým účinkom senzibilizácie nociceptora imunitnými faktormi je zvýšené uvoľňovanie neuropeptidov na periférnych zakončeniach, ktoré ďalej aktivujú imunitné bunky, čím sa indukuje pozitívna spätná väzba, ktorá riadi a uľahčuje zápal.
Kontrola vrodenej a adaptívnej imunity senzorickým nervovým systémom
V skorých fázach zápalu senzorické neuróny signalizujú žírnym bunkám a dendritickým bunkám, ktoré sú rezidentnými v tkanive, čo sú vrodené imunitné bunky dôležité pri spúšťaní imunitnej odpovede (obr. 2). Anatomické štúdie ukázali priamu apozíciu terminálov so žírnymi bunkami, ako aj s dendritickými bunkami a neuropeptidy uvoľnené z nociceptorov môžu indukovať degranuláciu alebo produkciu cytokínov v týchto bunkách 7, 9, 37. Táto interakcia hrá dôležitú úlohu pri alergických dýchacích cestách zápal a dermatitída 10�12.
Počas efektorovej fázy zápalu si imunitné bunky musia nájsť cestu k špecifickému miestu poranenia. Mnohé mediátory uvoľňované zo senzorických neurónov, neuropeptidy, chemokíny a glutamát, sú chemotaktické pre neutrofily, eozinofily, makrofágy a T-bunky a zvyšujú endoteliálnu adhéziu, ktorá uľahčuje navádzanie imunitných buniek 6, 38�41 (obr. 2). Okrem toho niektoré dôkazy naznačujú, že neuróny sa môžu priamo podieľať na efektorovej fáze, pretože samotné neuropeptidy môžu mať priame antimikrobiálne funkcie 42.
Neurónovo odvodené signálne molekuly môžu tiež riadiť typ zápalu tým, že prispievajú k diferenciácii alebo špecifikácii rôznych typov adaptívnych imunitných T buniek. Antigén je fagocytovaný a spracovaný bunkami vrodenej imunity, ktoré potom migrujú do najbližšej lymfatickej uzliny a prezentujú antigénny peptid natívnym T bunkám. V závislosti od typu antigénu, kostimulačných molekúl na vrodenej imunitnej bunke a kombinácií špecifických cytokínov dozrievajú natívne T bunky do špecifických podtypov, ktoré najlepšie slúžia zápalovému úsiliu odstrániť patogénny stimul. CD4 T bunky alebo T pomocné (Th) bunky možno rozdeliť do štyroch základných skupín, Th1, Th2, Th17 a T regulačné bunky (Treg). Th1 bunky sa podieľajú hlavne na regulácii imunitných odpovedí na intracelulárne mikroorganizmy a orgánovo špecifické autoimunitné ochorenia; Th2 sú kritické pre imunitu proti extracelulárnym patogénom, ako sú helminty, a sú zodpovedné za alergické zápalové ochorenia; Th17 bunky hrajú ústrednú úlohu pri ochrane proti mikrobiálnym výzvam, ako sú extracelulárne baktérie a huby; Treg bunky sa podieľajú na udržiavaní vlastnej tolerancie a regulácii imunitných reakcií. Zdá sa, že tento proces dozrievania T buniek je silne ovplyvnený senzorickými neurónovými mediátormi. Neuropeptidy, ako sú CGRP a VIP, môžu priviesť dendritické bunky k imunite typu Th2 a znížiť imunitu typu Th1 podporou produkcie určitých cytokínov a inhibíciou iných, ako aj znížením alebo zvýšením migrácie dendritických buniek do lokálnych lymfatických uzlín 8 , 10, 43. Senzorické neuróny tiež značne prispievajú k alergickému (hlavne Th2 riadenému) zápalu 17. Okrem regulácie Th1 a Th2 buniek môžu iné neuropeptidy, ako je SP a Hemokinín-1, poháňať zápalovú odpoveď viac smerom k Th17 alebo Treg 44, 45, čo znamená, že neuróny sa môžu tiež podieľať na regulácii zápalového riešenia. Pri imunopatologických stavoch, ako je kolitída a psoriáza, môže blokáda neuronálnych mediátorov, ako je látka P, významne tlmiť poškodenie T-buniek a imunitne sprostredkované15�17, hoci antagonizácia jedného mediátora môže mať sama osebe len obmedzený účinok na neurogénny zápal.
Vzhľadom na to, že signálne molekuly uvoľňované z periférnych senzorických nervových vlákien regulujú nielen malé krvné cievy, ale aj chemotaxiu, navádzanie, dozrievanie a aktiváciu imunitných buniek, začína byť jasné, že neuroimunitné interakcie sú oveľa zložitejšie, ako sa doteraz predpokladalo (obr. 2). Okrem toho je celkom možné, že to nie sú jednotlivé nervové mediátory, ale skôr špecifické kombinácie signálnych molekúl uvoľnených z nociceptorov, ktoré ovplyvňujú rôzne štádiá a typy imunitných reakcií.
Autonómna reflexná kontrola imunity
Zdá sa, že významná je aj úloha „reflexného okruhu“ cholinergného autonómneho nervového systému pri regulácii periférnych imunitných reakcií 46. Vagus je hlavný parasympatický nerv spájajúci mozgový kmeň s viscerálnymi orgánmi. Práca Kevina Traceyho a ďalších poukazuje na silné generalizované protizápalové reakcie pri septickom šoku a endotoxémii, ktoré sú spúšťané eferentnou aktivitou vagového nervu, ktorá vedie k potlačeniu periférnych makrofágov 47�49. Vagus aktivuje periférne adrenergné neuróny celiakálneho ganglia inervujúce slezinu, čo vedie k následnému uvoľňovaniu acetylcholínu, ktorý sa viaže na alfa-7 nikotínové receptory na makrofágoch v slezine a gastrointestinálnom trakte. To vyvoláva aktiváciu signálnej dráhy JAK2/STAT3 SOCS3, ktorá silne potláča transkripciu TNF-alfa 47. Adrenergný celiakálny ganglion tiež priamo komunikuje s podskupinou pamäťových T buniek produkujúcich acetylcholín, ktoré potláčajú zápalové makrofágy 48.
Invariantné prirodzené zabíjačské T bunky (iNKT) sú špecializovanou podskupinou T buniek, ktoré rozpoznávajú mikrobiálne lipidy v kontexte CD1d namiesto peptidových antigénov. NKT bunky sú kľúčovou populáciou lymfocytov zapojenou do boja proti infekčným patogénom a regulácie systémovej imunity. Bunky NKT sídlia a prechádzajú hlavne cez vaskulatúru a sínusoidy sleziny a pečene. Sympatické beta-adrenergné nervy v pečeni priamo signalizujú moduláciu aktivity NKT buniek 50. Napríklad počas myšacieho modelu mŕtvice (MCAO) bola mobilita NKT buniek pečene viditeľne potlačená, čo bolo zvrátené sympatickou denerváciou alebo beta-adrenergnými antagonistami. Okrem toho táto imunosupresívna aktivita noradrenergných neurónov na NKT bunkách viedla k zvýšeniu systémovej infekcie a poranenia pľúc. Preto eferentné signály z autonómnych neurónov môžu sprostredkovať silnú imunosupresiu.
Pohľad Dr. Alexa Jimeneza
Neurogénny zápal je lokálna zápalová reakcia generovaná nervovým systémom. Predpokladá sa, že hrá základnú úlohu v patogenéze rôznych zdravotných problémov, vrátane migrény, psoriázy, astmy, fibromyalgie, ekzému, rosacey, dystónie a mnohonásobnej chemickej citlivosti. Hoci bol neurogénny zápal spojený s periférnym nervovým systémom rozsiahlo skúmaný, koncept neurogénneho zápalu v centrálnom nervovom systéme ešte potrebuje ďalší výskum. Podľa niekoľkých výskumných štúdií sa však predpokladá, že nedostatok horčíka je hlavnou príčinou neurogénneho zápalu. Nasledujúci článok demonštruje prehľad mechanizmov neurogénneho zápalu v nervovom systéme, ktorý môže zdravotníckym pracovníkom pomôcť určiť najlepší liečebný prístup k starostlivosti o rôzne zdravotné problémy spojené s nervovým systémom.
Závery
Aké sú konkrétne úlohy somatosenzorického a autonómneho nervového systému pri regulácii zápalu a imunitného systému (obr. 4)? Aktivácia nociceptorov vedie k lokálnym axónovým reflexom, ktoré lokálne získavajú a aktivujú imunitné bunky, a preto sú hlavne prozápalové a priestorovo obmedzené. Na rozdiel od toho autonómna stimulácia vedie k systémovej imunosupresii ovplyvnením skupín imunitných buniek v pečeni a slezine. Mechanizmy aferentnej signalizácie na periférii vedúce k spusteniu imunosupresívneho vagového cholinergného reflexného okruhu nie sú dostatočne známe. Avšak 80 – 90 % vagových vlákien sú primárne aferentné senzorické vlákna, a preto signály z vnútorností, z ktorých mnohé sú potenciálne poháňané imunitnými bunkami, môžu viesť k aktivácii interneurónov v mozgovom kmeni a cez ne k výstupu v eferentných vagových vláknach 46.
Obrázok 4: Senzorické a autonómne nervové systémy modulujú lokálne a systémové imunitné reakcie. Nociceptory inervujúce povrchy epitelu (napr. kožu a pľúca) indukujú lokalizované zápalové reakcie, aktivujú žírne a dendritické bunky. Pri alergickom zápale dýchacích ciest, dermatitíde a reumatoidnej artritíde zohrávajú nociceptorové neuróny úlohu pri riadení zápalu. Naproti tomu autonómne okruhy inervujúce viscerálne orgány (napr. slezinu a pečeň) regulujú systémové imunitné reakcie blokovaním aktivácie makrofágov a NKT buniek. Pri mŕtvici a septickej endotoxémii hrajú tieto neuróny imunosupresívnu úlohu.
Typicky je časový priebeh a povaha zápalu, či už počas infekcie, alergických reakcií alebo autoimunitných patológií, definovaná kategóriami zahrnutých imunitných buniek. Bude dôležité vedieť, aké rôzne typy imunitných buniek sú regulované senzorickými a autonómnymi signálmi. Systematické hodnotenie toho, aké mediátory sa môžu uvoľňovať z nociceptorov a autonómnych neurónov a expresia receptorov pre ne rôznymi vrodenými a adaptívnymi imunitnými bunkami, môže pomôcť vyriešiť túto otázku.
Počas evolúcie sa vyvinuli podobné molekulárne dráhy na detekciu nebezpečenstva pre vrodenú imunitu aj nocicepciu, aj keď bunky majú úplne odlišné vývojové línie. Zatiaľ čo PRR a škodlivé iónové kanály riadené ligandom študujú imunológovia a neurobiológovia oddelene, hranica medzi týmito dvoma oblasťami je čoraz nejasnejšia. Počas poškodenia tkaniva a patogénnej infekcie uvoľnenie nebezpečných signálov pravdepodobne povedie ku koordinovanej aktivácii periférnych neurónov a imunitných buniek s komplexnou obojsmernou komunikáciou a integrovanou obranou hostiteľa. Anatomické umiestnenie nociceptorov na rozhraní s prostredím, rýchlosť nervovej transdukcie a ich schopnosť uvoľňovať silné koktaily imunitne pôsobiacich mediátorov umožňuje periférnemu nervovému systému aktívne modulovať vrodenú imunitnú reakciu a koordinovať adaptívnu imunitu. Naopak, nociceptory sú vysoko citlivé na imunitné mediátory, ktoré aktivujú a senzibilizujú neuróny. Neurogénne a imunitne sprostredkované zápaly preto nie sú nezávislé jednotky, ale pôsobia spoločne ako zariadenia včasného varovania. Avšak periférny nervový systém tiež hrá dôležitú úlohu v patofyziológii, a možno etiológie, mnohých imunitných ochorení, ako je astma, psoriáza alebo kolitída, pretože jeho schopnosť aktivovať imunitný systém môže zosilniť patologický zápal 15�17. Preto môže byť potrebné, aby liečba imunitných porúch zahŕňala zacielenie nociceptorov, ako aj imunitných buniek.
Poďakovanie
Ďakujeme NIH za podporu (2R37NS039518).
Záverom možno povedať,Pochopenie úlohy neurogénneho zápalu, pokiaľ ide o obranu hostiteľa a imunopatológiu, je nevyhnutné na určenie správneho liečebného prístupu pre rôzne zdravotné problémy nervového systému. Pri pohľade na interakcie periférnych neurónov s imunitnými bunkami môžu zdravotnícki pracovníci pokročiť v terapeutických prístupoch, aby ďalej pomohli zvýšiť obranu hostiteľa, ako aj potlačiť imunopatológiu. Účelom vyššie uvedeného výrobku je pomôcť pacientom pochopiť klinickú neurophysiológiu neuropatie, okrem iných zdravotných problémov zranenia nervu. Informácie s odkazom na Národné centrum pre biotechnologické informácie (NCBI). Rozsah našich informácií je obmedzený na chiropraktiku, ako aj na poranenia a stavy chrbtice. Ak chcete prediskutovať túto tému, neváhajte sa opýtať Dr. Jimeneza alebo nás kontaktujte na adrese�915-850-0900 .
Kurátorom je Dr. Alex Jimenez
Ďalšie témy: Bolesť chrbta
Bolesti chrbta je jednou z najčastejších príčin invalidity a vymeškaných dní v práci na celom svete. Bolesti chrbta sa v skutočnosti pripisujú ako druhý najčastejší dôvod návštevy lekára, prevyšujú ju len infekcie horných dýchacích ciest. Približne 80 percent populácie aspoň raz za život zažije nejaký typ bolesti chrbta. Chrbtica je zložitá štruktúra pozostávajúca z kostí, kĺbov, väzov a svalov, okrem iného z mäkkých tkanív. Z tohto dôvodu môžu zranenia a/alebo zhoršené stavy, ako napr herné disky, môže nakoniec viesť k príznakom bolesti chrbta. Športové zranenia alebo zranenia pri automobilových nehodách sú často najčastejšou príčinou bolesti chrbta, niekedy však môžu mať aj tie najjednoduchšie pohyby bolestivé následky. Našťastie alternatívne možnosti liečby, ako je chiropraktická starostlivosť, môžu pomôcť zmierniť bolesť chrbta pomocou úprav chrbtice a manuálnych manipulácií, čo v konečnom dôsledku zlepšuje úľavu od bolesti.
1.�Sauer Sk, Reeh Pw, Boj GM. Škodlivé teplom indukované uvoľňovanie CGRP z axónov sedacieho nervu potkanov in vitro.�Eur J Neurosci.�2001;14:1203[PubMed]
2.�Edvinsson L, Ekman R, Jansen I, McCulloch J, Uddman R. Peptid súvisiaci s génom kalcitonínu a cerebrálne krvné cievy: distribúcia a vazomotorické účinky.�J Cereb Blood Flow Metab.�1987;7:720[PubMed]
3.�McCormack DG, Mak JC, Coupe MO, Barnes PJ. Vazodilatácia peptidov súvisiacich s kalcitonínovým génom ľudských pľúcnych ciev.�J Appl Physiol.�1989;67:1265[PubMed]
4.�Saria A. Látka P v senzorických nervových vláknach prispieva k rozvoju edému zadnej labky potkana po tepelnom poranení.�Br J Pharmacol.�1984;82:217[Článok bez PMC]�[PubMed]
5.�Brain SD, Williams TJ. Interakcie medzi tachykinínmi a peptidom súvisiacim s génom kalcitonínu vedú k modulácii tvorby edému a prietoku krvi v koži potkanov.�Br J Pharmacol.�1989;97:77.[Článok bez PMC]�[PubMed]
6.�Fryer AD, a kol. Neurónový eotaxín a účinky antagonistu CCR3 na hyperreaktivitu dýchacích ciest a dysfunkciu M2 receptora.�J Clin Invest.�2006;116:228[Článok bez PMC]�[PubMed]
7.�Ansel JC, Brown JR, Payan DG, Brown MA. Látka P selektívne aktivuje expresiu génu TNF-alfa v myších žírnych bunkách.�J Immunol.�1993;150:4478[PubMed]
8.�Ding W, Stohl LL, Wagner JA, Granstein RD. Peptid súvisiaci s génom kalcitonínu privádza Langerhansove bunky smerom k imunite typu Th2.�J Immunol.�2008;181:6020[Článok bez PMC]�[PubMed]
9.�Hosoi J, a kol. Regulácia funkcie Langerhansových buniek nervami obsahujúcimi peptid súvisiaci s génom kalcitonínu.�Príroda.�1993;363:159[PubMed]
10.�Mikami N, a kol. Peptid súvisiaci s génom kalcitonínu je dôležitým regulátorom kožnej imunity: účinok na dendritické bunky a funkcie T buniek.�J Immunol.�2011;186:6886[PubMed]
11.�Rochlitzer S, a kol. Neuropeptidový peptid súvisiaci s génom kalcitonínu ovplyvňuje alergický zápal dýchacích ciest moduláciou funkcie dendritických buniek.�Clin Exp Allergy.�2011;41:1609[PubMed]
12.�Cyphert JM a kol. Spolupráca medzi mastocytmi a neurónmi je nevyhnutná pre antigénom sprostredkovanú bronchokonstrikciu.�J Immunol.�2009;182:7430[Článok bez PMC]�[PubMed]
13.�Levine JD a kol. Intraneuronálna látka P prispieva k závažnosti experimentálnej artritídy.�Veda.�1984;226:547[PubMed]
14.�Levine JD, Khasar SG, Green PG. Neurogénny zápal a artritída.�Ann NY Acad Sci.�2006;1069:155[PubMed]
15.�Engel MA a kol. TRPA1 a látka P sprostredkúvajú kolitídu u myší.�Gastroenterológia.�2011;141:1346[PubMed]
16.�Ostrowski SM, Belkadi A, Loyd CM, Diaconu D, Ward NL. Kožná denervácia kože psoriasiformných myší zlepšuje akantózu a zápal senzoricky závislým spôsobom od neuropeptidov.�J Invest Dermatol.�2011;131:1530[Článok bez PMC]�[PubMed]
17.�Caceres AI a kol. Senzorický neurónový iónový kanál nevyhnutný pre zápal dýchacích ciest a hyperreaktivitu pri astme.�Proc Natl Acad Sci USA A.�2009;106:9099[Článok bez PMC]�[PubMed]
18.�Caterina MJ a kol. Zhoršená nocicepcia a vnímanie bolesti u myší, ktorým chýba kapsaicínový receptor.�Veda.�2000;288:306[PubMed]
19.�Bessac BF a kol. Antagonisty ankyrínu 1 s prechodným receptorovým potenciálom blokujú škodlivé účinky toxických priemyselných izokyanátov a slzných plynov.�FASEB J.�2009;23:1102[Článok bez PMC]�[PubMed]
20.�Cruz-Orengo L, a kol. Kožná nocicepcia vyvolaná 15-delta PGJ2 prostredníctvom aktivácie iónového kanála TrPA1.�Mol Pain.�2008;4:30[Článok bez PMC]�[PubMed]
21.�Trevisani M, a kol. 4-Hydroxynonenal, endogénny aldehyd, spôsobuje bolesť a neurogénny zápal prostredníctvom aktivácie dráždivého receptora TRPA1.�Proc Natl Acad Sci USA A.�2007;104:13519[Článok bez PMC]�[PubMed]
22.�Janeway CA, Jr., Medzhitov R. Úvod: Úloha vrodenej imunity v adaptívnej imunitnej odpovedi.�Semin Immunol.�1998;10:349[PubMed]
23.�Matzinger P. Vrodený pocit nebezpečenstva.�Ann NY Acad Sci.�2002;961:341[PubMed]
24.�Bianchi ME. DAMP, PAMP a alarmíny: všetko, čo potrebujeme vedieť o nebezpečenstve.�J Leukoc Biol.�2007;81:1[PubMed]
25.�Liu T, Xu ZZ, Park CK, Berta T, Ji RR. Toll-like receptor 7 sprostredkuje pruritus.�Nat Neurosci.�2010;13:1460[Článok bez PMC]�[PubMed]
26.�Diogenes A, Ferraz CC, Akopian AN, Henry MA, Hargreaves KM. LPS senzibilizuje TRPV1 prostredníctvom aktivácie TLR4 v senzorických neurónoch trigeminu.�J Dent Res.�2011;90:759[PubMed]
27.�Qi J, a kol. Bolestivé dráhy vyvolané TLR stimuláciou neurónov ganglií dorzálnych koreňov.�J Immunol.�2011;186:6417[Článok bez PMC]�[PubMed]
28.�Cockayne DA a kol. Hyporeflexia močového mechúra a znížené správanie súvisiace s bolesťou u myší s deficitom P2X3.�Príroda.�2000;407:1011[PubMed]
29.�Mariathasan S, a kol. Kryopyrín aktivuje zápal v reakcii na toxíny a ATP.�Príroda.�2006;440:228[PubMed]
30.�Souslova V, et al. Deficity teplého kódovania a aberantná zápalová bolesť u myší bez receptorov P2X3.�Príroda.�2000;407:1015[PubMed]
31.�de Rivero Vaccari JP, Lotocki G, Marcillo AE, Dietrich WD, Keane RW. Molekulárna platforma v neurónoch reguluje zápal po poranení miechy.�J Neurosci.�2008;28:3404[PubMed]
32.�Link TM a kol. TRPV2 má kľúčovú úlohu pri viazaní makrofágových častíc a fagocytóze.�Nat Immunol.�2010;11:232[Článok bez PMC]�[PubMed]
33.�Turner H, del Carmen KA, Stokes A. Prepojenie medzi kanálmi TRPV a funkciou žírnych buniek.�Handb Exp Pharmacol.�2007: 457�471.�[PubMed]
34.�Binshtok AM, a kol. Nociceptory sú senzory interleukínu-1beta.�J Neurosci.�2008;28:14062.[Článok bez PMC]�[PubMed]
35.�Zhang XC, Kainz V, Burstein R, Levy D. Tumor nekrotizujúci faktor-alfa indukuje senzibilizáciu meningeálnych nociceptorov sprostredkovanú lokálnym pôsobením COX a p38 MAP kinázy.�Bolesť.�2011;152:140.[Článok bez PMC]�[PubMed]
36.�Samad TA a kol. Interleukínom-1beta sprostredkovaná indukcia Cox-2 v CNS prispieva k hypersenzitivite zápalovej bolesti.�Príroda.�2001;410:471[PubMed]
37.�Veres TZ a kol. Priestorové interakcie medzi dendritickými bunkami a zmyslovými nervami pri alergickom zápale dýchacích ciest.�AM J J Respir Cell Mol Biol.�2007;37:553[PubMed]
38.�Smith CH, Barker JN, Morris RW, MacDonald DM, Lee TH. Neuropeptidy indukujú rýchlu expresiu adhéznych molekúl endotelových buniek a vyvolávajú granulocytovú infiltráciu v ľudskej koži.�J Immunol.�1993;151:3274[PubMed]
39.�Dunzendorfer S, Meierhofer C, Wiedermann CJ. Signalizácia pri migrácii ľudských eozinofilov indukovanej neuropeptidmi.�J Leukoc Biol.�1998;64:828[PubMed]
40.�Ganor Y, Besser M, Ben-Zakay N, Unger T, Levite M. Ľudské T bunky exprimujú funkčný ionotropný glutamátový receptor GluR3 a glutamát samotný spúšťa integrínom sprostredkovanú adhéziu k laminínu a fibronektínu a chemotaktickú migráciu.�J Immunol.�2003;170:4362[PubMed]
41.�CzePielewski Rs a kol. Peptidový receptor uvoľňujúci gastrín (GRPR) sprostredkúva chemotaxiu v neutrofiloch.�Proc Natl Acad Sci USA A.�2011;109:547[Článok bez PMC]�[PubMed]
42.�Brogden KA, Guthmiller JM, Salzet M, Zasloff M. Nervový systém a vrodená imunita: Neuropeptidové pripojenie.�Nat Immunol.�2005;6:558[PubMed]
43.�Jimeno R, a kol. Účinok VIP na rovnováhu medzi cytokínmi a hlavnými regulátormi aktivovaných pomocných T buniek.�Immunol Cell Biol.�2011;90:178[PubMed]
44.�Razavi R, a kol. Senzorické neuróny TRPV1+ kontrolujú stres beta buniek a zápal ostrovčekov pri autoimunitnom diabete.�Bunka.�2006;127:1123[PubMed]
45.�Cunin P, a kol. Tachykinínová látka P a hemokinín-1 podporujú tvorbu ľudských pamäťových Th17 buniek indukciou expresie IL-1beta, IL-23 a TNF podobnej 1A monocytmi.�J Immunol.�2011;186:4175[PubMed]
46.�Andersson U, Tracey KJ. Reflexné princípy imunologickej homeostázy.�Annu Rev Immunol.�2011[Článok bez PMC]�[PubMed]
47.�de Jonge WJ a kol. Stimulácia nervu vagus zoslabuje aktiváciu makrofágov aktiváciou signálnej dráhy Jak2-STAT3.�Nat Immunol.�2005;6:844[PubMed]
48.�Rosas-Ballina M, a kol. Acetylcholín syntetizujúce T bunky prenášajú nervové signály v okruhu nervu vagus.�Veda.�2011;334:98[Článok bez PMC]�[PubMed]
49.�Wang H a kol. Podjednotka nikotínového acetylcholínového receptora alfa7 je základným regulátorom zápalu.�Príroda.�2003;421:384[PubMed]
50.�Wong CH, Jenne CN, Lee WY, Leger C, Kubes P. Funkčná inervácia pečeňových iNKT buniek je po mŕtvici imunosupresívna.�Veda.�2011;334:101[PubMed]
Nástroj Find A Practitioner od IFM je najväčšou sieťou odporúčaní vo funkčnej medicíne, ktorá bola vytvorená s cieľom pomôcť pacientom nájsť praktických lekárov kdekoľvek na svete. IFM Certified Practitioners sú vo výsledkoch vyhľadávania uvedení na prvom mieste vzhľadom na ich rozsiahle vzdelanie v oblasti funkčnej medicíny



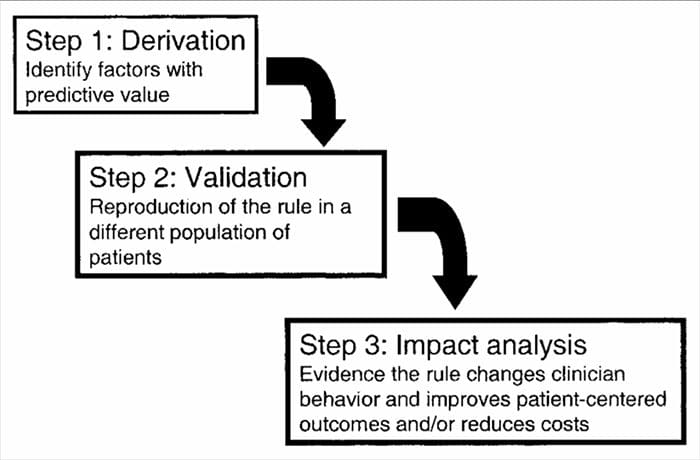




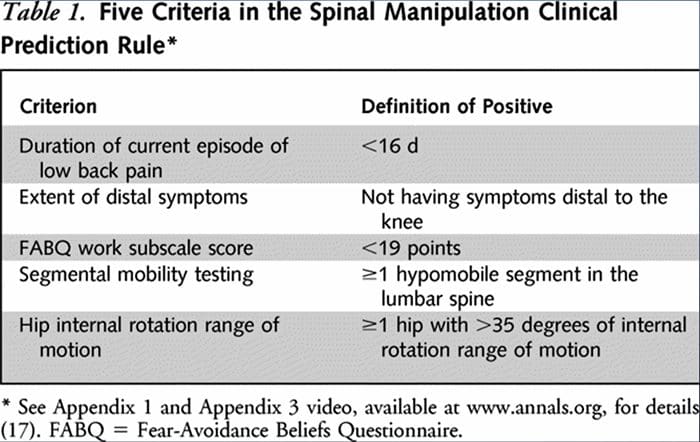
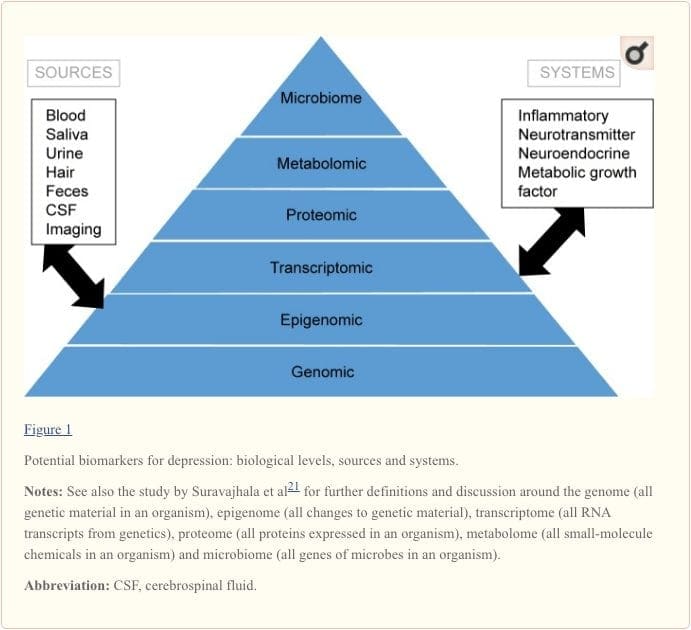





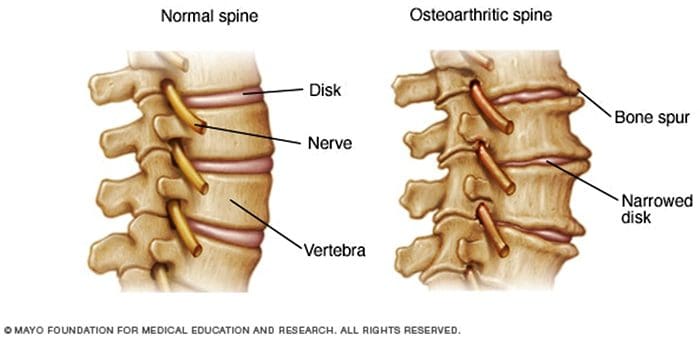
 abstraktné
abstraktné

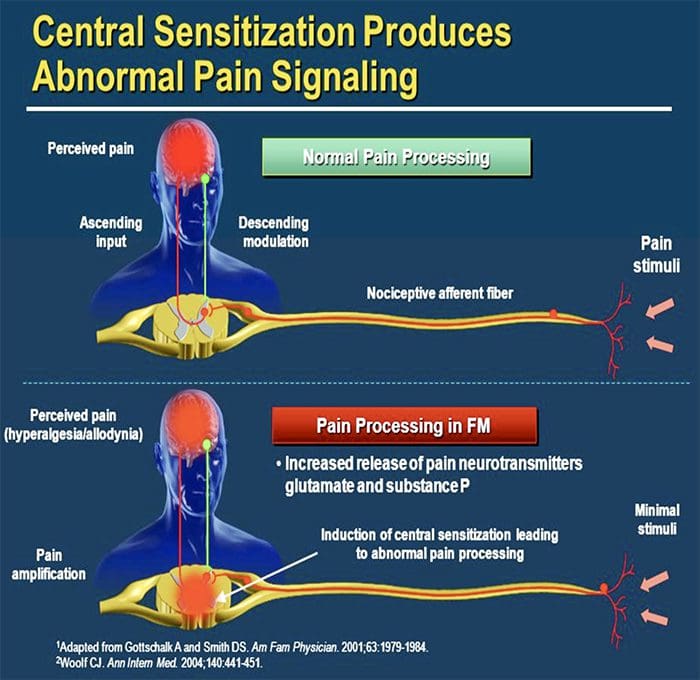


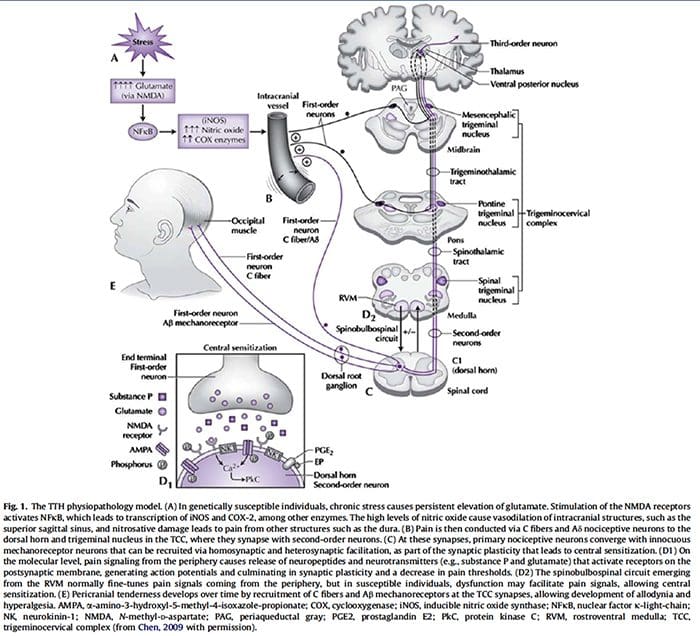

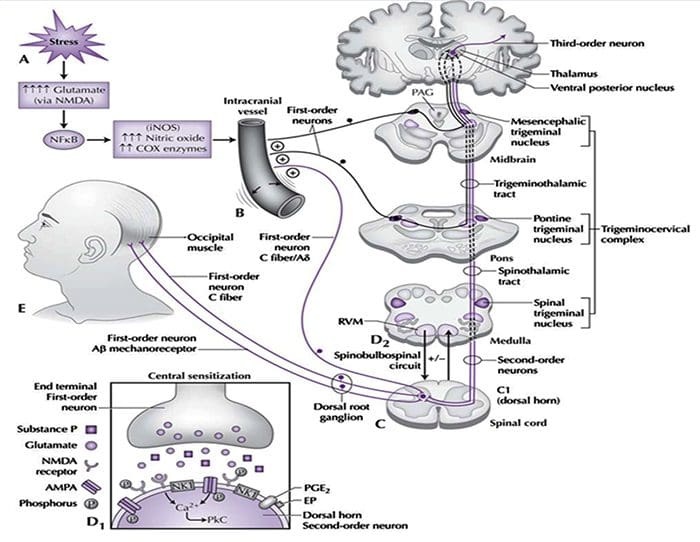




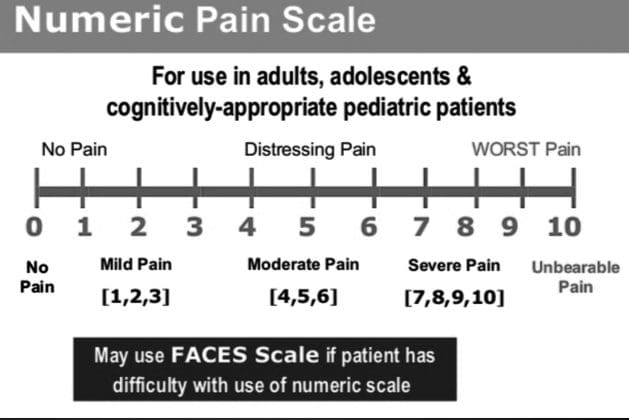










 Referencie:
Referencie:
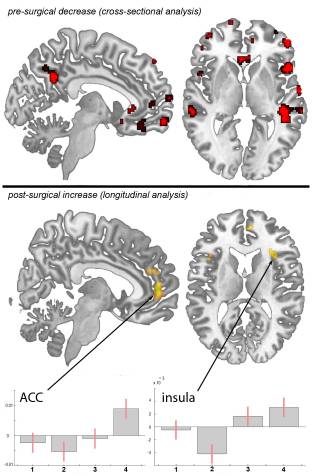
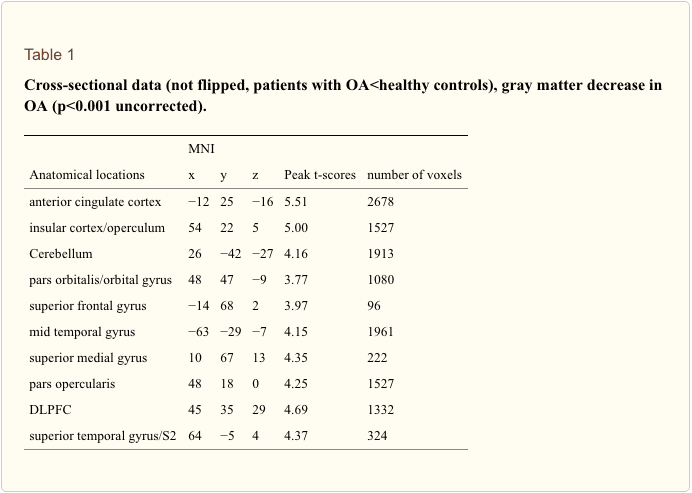
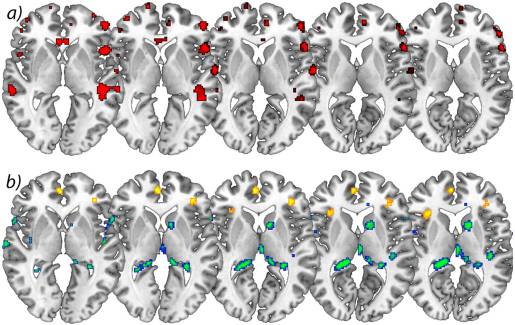
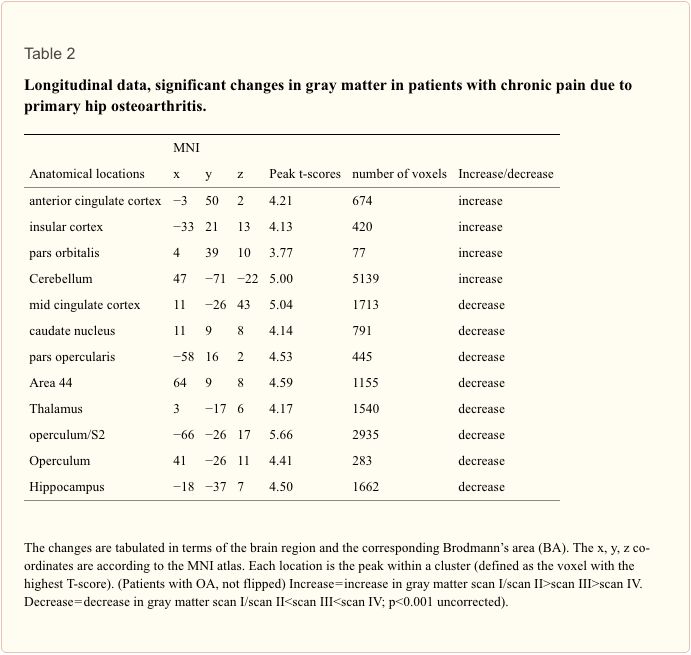
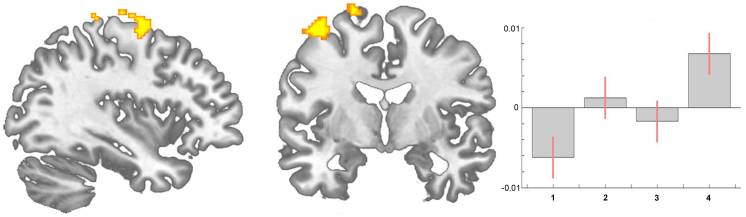
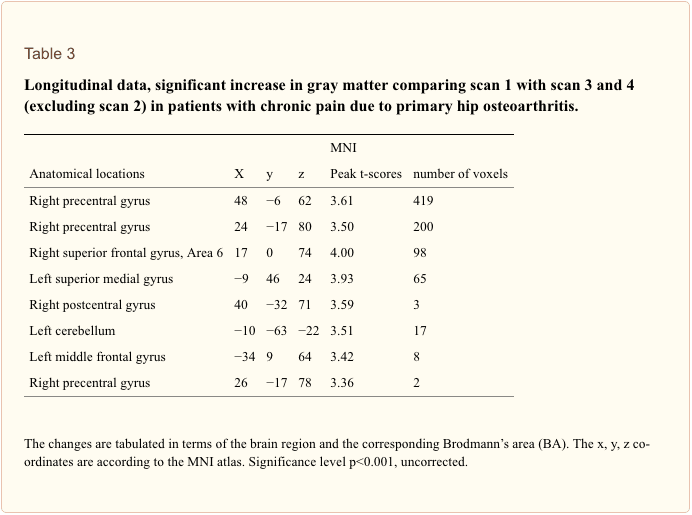

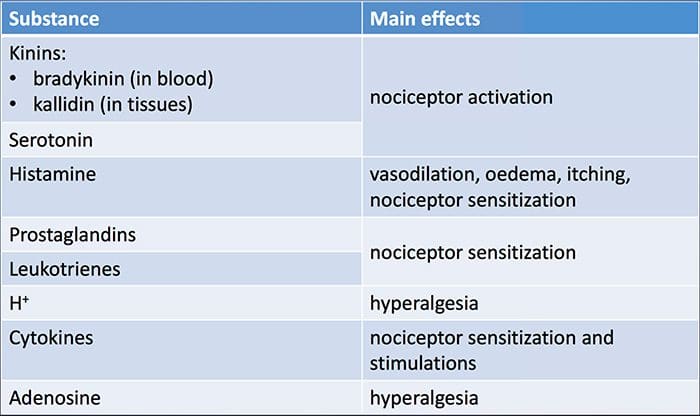
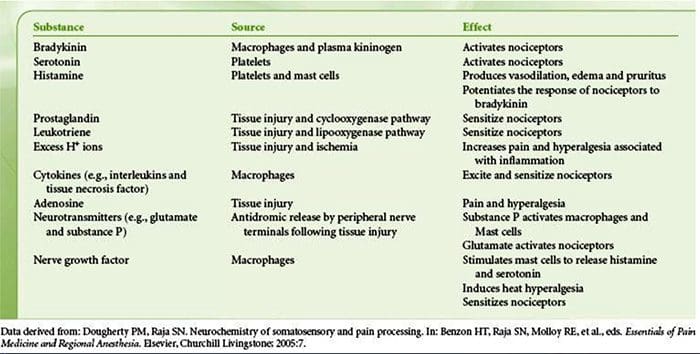
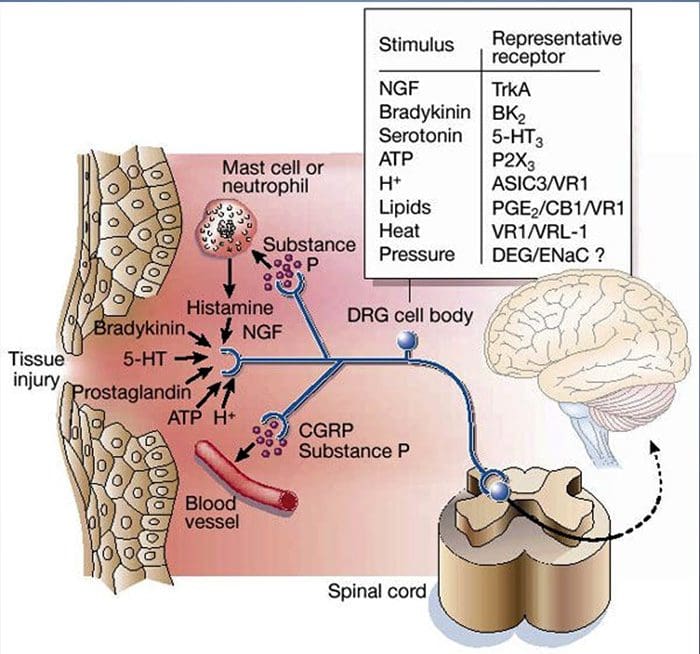
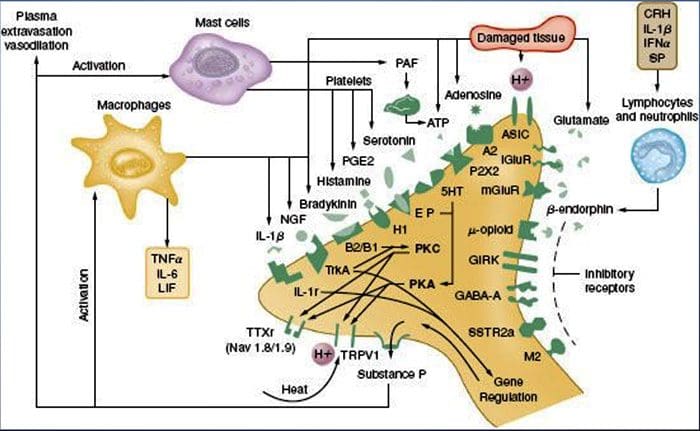 Prečo ma bolí rameno? Prehľad neuroanatomického a biochemického základu bolesti ramena
Prečo ma bolí rameno? Prehľad neuroanatomického a biochemického základu bolesti ramena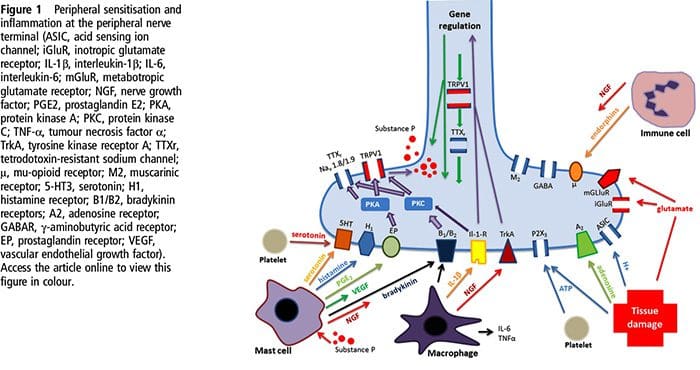 NGF a receptor katiónového kanála s prechodným potenciálom receptora V člen 1 (TRPV1) majú symbiotický vzťah, pokiaľ ide o zápal a senzibilizáciu nociceptora. Cytokíny produkované v zapálenom tkanive vedú k zvýšeniu produkcie NGF.19 NGF stimuluje uvoľňovanie histamínu a serotonínu (5-HT3) žírnymi bunkami a tiež senzibilizuje nociceptory, čo môže zmeniť vlastnosti A? vlákna tak, že väčšia časť sa stáva nociceptívnou. Receptor TRPV1 je prítomný v subpopulácii primárnych aferentných vlákien a je aktivovaný kapsaicínom, teplom a protónmi. Receptor TRPV1 je syntetizovaný v bunkovom tele aferentného vlákna a je transportovaný do periférnych aj centrálnych zakončení, kde prispieva k senzitivite nociceptívnych aferentov. Zápal vedie k periférnej produkcii NGF, ktorý sa potom naviaže na receptor tyrozínkinázy typu 1 na zakončeniach nociceptora, NGF je potom transportovaný do bunkového tela, kde vedie k upregulácii transkripcie TRPV1 a následne k zvýšeniu citlivosti nociceptora.19 20 NGF a iné zápalové mediátory tiež senzibilizujú TRPV1 prostredníctvom rôznorodého radu sekundárnych mediátorových dráh. Mnoho ďalších receptorov vrátane cholinergných receptorov, receptorov kyseliny a-aminomaslovej (GABA) a somatostatínových receptorov sa tiež považuje za zapojené do periférnej citlivosti nociceptorov.
NGF a receptor katiónového kanála s prechodným potenciálom receptora V člen 1 (TRPV1) majú symbiotický vzťah, pokiaľ ide o zápal a senzibilizáciu nociceptora. Cytokíny produkované v zapálenom tkanive vedú k zvýšeniu produkcie NGF.19 NGF stimuluje uvoľňovanie histamínu a serotonínu (5-HT3) žírnymi bunkami a tiež senzibilizuje nociceptory, čo môže zmeniť vlastnosti A? vlákna tak, že väčšia časť sa stáva nociceptívnou. Receptor TRPV1 je prítomný v subpopulácii primárnych aferentných vlákien a je aktivovaný kapsaicínom, teplom a protónmi. Receptor TRPV1 je syntetizovaný v bunkovom tele aferentného vlákna a je transportovaný do periférnych aj centrálnych zakončení, kde prispieva k senzitivite nociceptívnych aferentov. Zápal vedie k periférnej produkcii NGF, ktorý sa potom naviaže na receptor tyrozínkinázy typu 1 na zakončeniach nociceptora, NGF je potom transportovaný do bunkového tela, kde vedie k upregulácii transkripcie TRPV1 a následne k zvýšeniu citlivosti nociceptora.19 20 NGF a iné zápalové mediátory tiež senzibilizujú TRPV1 prostredníctvom rôznorodého radu sekundárnych mediátorových dráh. Mnoho ďalších receptorov vrátane cholinergných receptorov, receptorov kyseliny a-aminomaslovej (GABA) a somatostatínových receptorov sa tiež považuje za zapojené do periférnej citlivosti nociceptorov.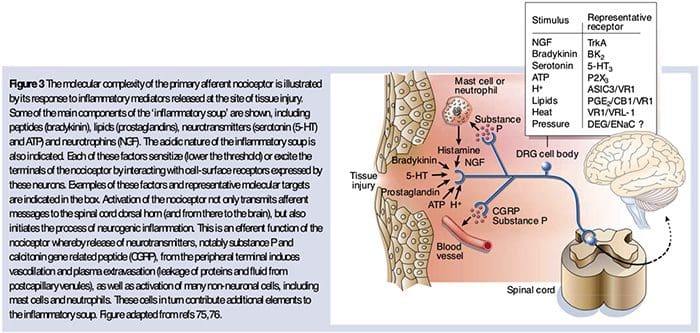 Neurochémia nociceptorov
Neurochémia nociceptorov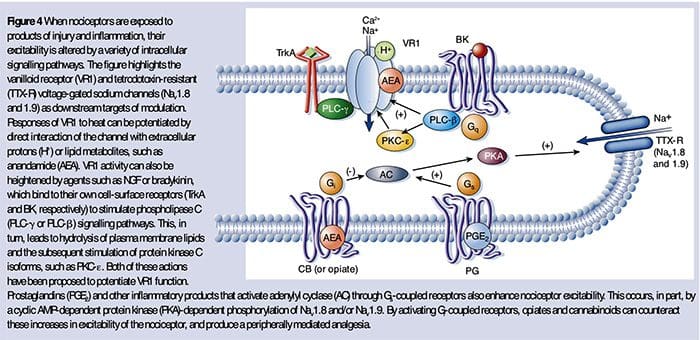 Bunkové a molekulárne mechanizmy bolesti
Bunkové a molekulárne mechanizmy bolesti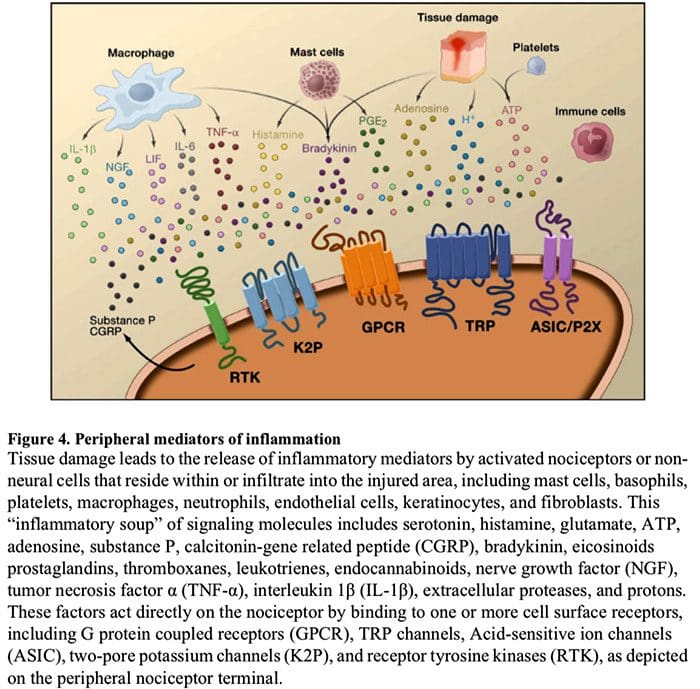
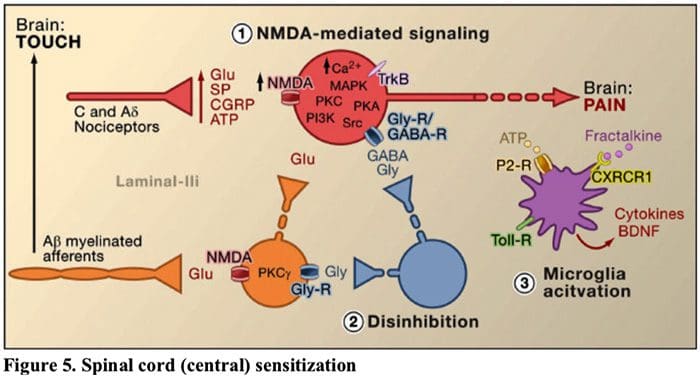 Obrázok 5. Miechová (centrálna) senzibilizácia
Obrázok 5. Miechová (centrálna) senzibilizácia