Back Clinic Podpora klinickej neurofyziológie. El Paso, TX. Hovorí chiropraktik Dr. Alexander Jimenez klinická neurofyziológia. Dr. Jimenez bude skúmať klinický význam a funkčné aktivity periférnych nervových vlákien, miechy, mozgového kmeňa a mozgu v kontexte viscerálnych a muskuloskeletálnych porúch. Pacienti získajú pokročilé znalosti o anatómii, genetike, biochémii a fyziológii bolesti vo vzťahu k rôznym klinickým syndrómom. Bude zahrnutá nutričná biochémia súvisiaca s nocicepciou a bolesťou. A dôraz sa bude klásť na implementáciu týchto informácií do terapeutických programov.
Náš tím je veľmi hrdý na to, že našim rodinám a zraneným pacientom prináša iba overené liečebné protokoly. Učením úplného holistického wellness ako životného štýlu meníme nielen životy našich pacientov, ale aj ich rodiny. Robíme to preto, aby sme mohli osloviť čo najviac obyvateľov El Paso, ktorí nás potrebujú, bez ohľadu na problémy s cenovou dostupnosťou. Pre odpovede na akékoľvek otázky, ktoré môžete mať, zavolajte Dr. Jimenezovi na číslo 915-850-0900.
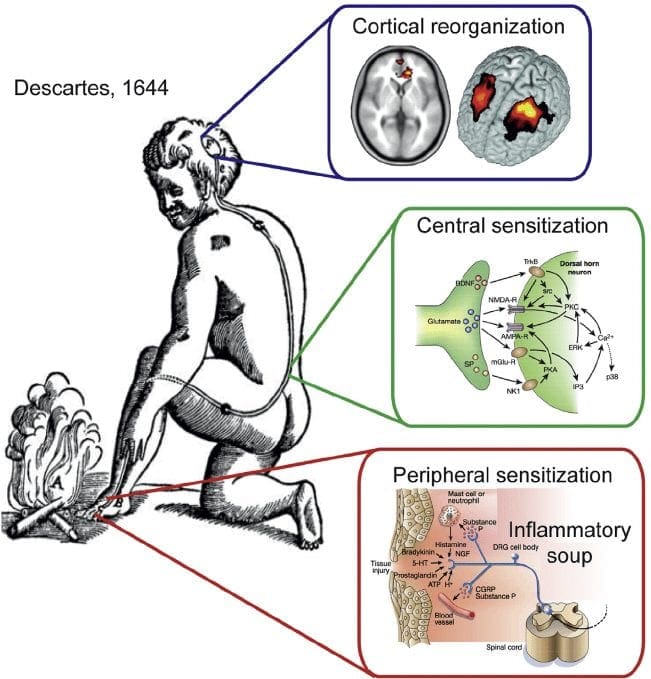
Neuropatická bolesť je komplexný, chronický bolestivý stav, ktorý je vo všeobecnosti sprevádzaný poranením mäkkých tkanív. Neuropatická bolesť je bežná v klinickej praxi a tiež predstavuje výzvu pre pacientov aj lekárov. Pri neuropatickej bolesti môžu byť samotné nervové vlákna buď poškodené, nefunkčné alebo zranené. Neuropatická bolesť je výsledkom poškodenia periférneho alebo centrálneho nervového systému v dôsledku traumy alebo choroby, kde sa lézia môže vyskytnúť na akomkoľvek mieste. V dôsledku toho môžu tieto poškodené nervové vlákna vysielať nesprávne signály do iných centier bolesti. Účinok poranenia nervových vlákien spočíva v zmene neurálnej funkcie, a to ako v oblasti poranenia, tak aj v okolí poranenia. Klinické príznaky neuropatickej bolesti normálne zahŕňajú senzorické javy, ako je spontánna bolesť, parestézie a hyperalgézia.
Neuropatická bolesť, ako ju definuje International Association of the Study of Pain alebo IASP, je bolesť iniciovaná alebo spôsobená primárnou léziou alebo dysfunkciou nervového systému. Mohlo by to vyplynúť z poškodenia kdekoľvek pozdĺž neuraxie: periférny nervový systém, miechový alebo supraspinálny nervový systém. Medzi znaky, ktoré odlišujú neuropatickú bolesť od iných druhov bolesti, patrí bolesť a zmyslové znaky trvajúce aj po období zotavenia. U ľudí je charakterizovaná spontánnou bolesťou, alodýniou alebo zážitkom neškodlivej stimulácie ako bolestivá a kauzalgia alebo pretrvávajúca pálivá bolesť. Spontánna bolesť zahŕňa pocity pichania, pálenia, vystreľovania, bodania a záchvatovitej bolesti alebo bolesti podobné elektrickému šoku, často spojené s dysestéziami a parestéziami. Tieto vnemy menia nielen pacientov zmyslový aparát, ale aj pacientovu pohodu, náladu, pozornosť a myslenie. Neuropatická bolesť sa skladá z „negatívnych“ symptómov, ako je strata zmyslov a pocity brnenia, ako aj z „pozitívnych“ symptómov, ako sú parestézie, spontánna bolesť a zvýšený pocit bolesti.
Stavy často súvisiace s neuropatickou bolesťou možno rozdeliť do dvoch hlavných skupín: bolesť spôsobená poškodením centrálneho nervového systému a bolesť spôsobená poškodením periférneho nervového systému. Kortikálne a subkortikálne mozgové príhody, traumatické poranenia miechy, syringomyélia a syringobulbia, neuralgia trojklaného nervu a glosofaryngeálneho nervu, neoplastické a iné lézie zaberajúce priestor sú klinické stavy, ktoré patria do prvej skupiny. Nervové kompresívne alebo zachytené neuropatie, ischemická neuropatia, periférne polyneuropatie, plexopatie, kompresia nervových koreňov, bolesť po amputácii pahýľa a fantómové končatiny, postherpetická neuralgia a neuropatie súvisiace s rakovinou sú klinické stavy, ktoré patria do druhej skupiny.
Patofyziológia neuropatickej bolesti
Patofyziologické procesy a koncepty, ktoré sú základom neuropatickej bolesti, sú viaceré. Pred pokrytím týchto procesov je kritická kontrola bežných obvodov bolesti. Pravidelné obvody bolesti zahŕňajú aktiváciu nociceptora, tiež známeho ako receptor bolesti, v reakcii na bolestivú stimuláciu. Vlna depolarizácie je dodaná do neurónov prvého rádu spolu so sodíkovým prúdením cez sodíkové kanály a uvoľňovaním draslíka von. Neuróny končia v mozgovom kmeni v trigeminálnom jadre alebo v dorzálnom rohu miechy. Je to tu, kde znak otvára napäťovo riadené vápnikové kanály v presynaptickom termináli, čo umožňuje vápniku vstúpiť. Vápnik umožňuje uvoľňovanie glutamátu, excitačného neurotransmitera, do synaptickej oblasti. Glutamát sa viaže na NMDA receptory na neurónoch druhého rádu, čo spôsobuje depolarizáciu.
Tieto neuróny prechádzajú cez miechu a cestujú až do talamu, kde sa synapsujú s neurónmi tretieho rádu. Tie sa následne spájajú s limbickým systémom a mozgovou kôrou. Existuje aj inhibičná dráha, ktorá zabraňuje prenosu signálu bolesti z dorzálneho rohu. Antinociceptívne neuróny pochádzajú z mozgového kmeňa a cestujú po mieche, kde sa synapsiou s krátkymi interneurónmi v dorzálnom rohu uvoľňujú dopamín a noradrenalín. Interneuróny modulujú synapsiu medzi neurónom prvého rádu, ako aj neurónom druhého rádu uvoľňovaním kyseliny gama aminomaslovej alebo GABA, inhibičného neurotransmitera. V dôsledku toho je zastavenie bolesti výsledkom inhibície synapsií medzi neurónmi prvého a druhého rádu, zatiaľ čo zosilnenie bolesti môže byť výsledkom potlačenia inhibičných synaptických spojení.
Mechanizmus, ktorý je základom neuropatickej bolesti, však nie je taký jasný. Niekoľko štúdií na zvieratách odhalilo, že môže byť zapojených veľa mechanizmov. Treba si však uvedomiť, že to, čo platí pre stvorenia, nemusí vždy platiť pre ľudí. Neuróny prvého rádu môžu zvýšiť svoju streľbu, ak sú čiastočne poškodené a zvýšiť množstvo sodíkových kanálov. Ektopické výboje sú dôsledkom zvýšenej depolarizácie na určitých miestach vlákna, čo vedie k spontánnej bolesti a bolesti súvisiacej s pohybom. Inhibičné okruhy môžu byť znížené na úrovni dorzálneho rohu alebo mozgových kmeňových buniek, ako aj oboch, čo umožňuje impulzom bolesti cestovať bez odporu.
Okrem toho môže dôjsť k zmenám v centrálnom spracovaní bolesti, keď v dôsledku chronickej bolesti a užívania niektorých liekov a/alebo liekov môžu neuróny druhého a tretieho rádu vytvoriť „pamäť“ bolesti a stať sa senzibilizovanými. Potom je tu zvýšená citlivosť miechových neurónov a znížené aktivačné prahy. Ďalšia teória demonštruje koncept sympatikom udržiavanej neuropatickej bolesti. Táto predstava bola preukázaná analgéziou po sympatektómii zvierat a ľudí. Pri mnohých chronických neuropatických alebo zmiešaných somatických a neuropatických bolestivých stavoch však môže byť zapojená aj zmes mechanizmov. Medzi tieto výzvy v oblasti bolesti a oveľa viac, pokiaľ ide o neuropatickú bolesť, patrí schopnosť kontrolovať ju. Má to dve zložky: po prvé, hodnotenie kvality, intenzity a pokroku; a po druhé, správne diagnostikovať neuropatickú bolesť.
Existujú však niektoré diagnostické nástroje, ktoré môžu lekárom pomôcť pri hodnotení neuropatickej bolesti. Pre začiatok, štúdie nervovej vodivosti a senzoricky evokované potenciály môžu identifikovať a kvantifikovať rozsah poškodenia senzorických, ale nie nociceptívnych dráh monitorovaním neurofyziologických reakcií na elektrické stimuly. Okrem toho kvantitatívne senzorické testovanie stupňuje vnímanie v reakcii na vonkajšie podnety rôznej intenzity aplikáciou stimulácie na pokožku. Mechanická citlivosť na hmatové podnety sa meria pomocou špecializovaných nástrojov, ako sú von Freyove chĺpky, pichnutie špendlíkom s do seba zapadajúcimi ihlami, ako aj citlivosť na vibrácie spolu s vibrametrami a tepelná bolesť pomocou termod.
Je tiež mimoriadne dôležité vykonať komplexné neurologické vyšetrenie na identifikáciu motorických, senzorických a autonómnych dysfunkcií. Nakoniec existuje množstvo dotazníkov používaných na rozlíšenie neuropatickej bolesti od nociceptívnej bolesti. Niektoré z nich obsahujú iba otázky na pohovor (napr. Neuropatický dotazník a bolesť ID), zatiaľ čo iné obsahujú otázky na pohovor aj fyzikálne testy (napr. Leedsova stupnica na hodnotenie neuropatických symptómov a znakov) a presný nový nástroj, štandardizované hodnotenie Bolesť, ktorá kombinuje šesť otázok na pohovor a desať fyziologických hodnotení.
Spôsoby liečby neuropatickej bolesti
Farmakologické režimy sú zamerané na mechanizmy neuropatickej bolesti. Farmakologická aj nefarmakologická liečba však prináša úplnú alebo čiastočnú úľavu len asi polovici pacientov. Mnohé svedectvá založené na dôkazoch naznačujú používanie zmesí liekov a/alebo liekov, aby fungovali pre čo najviac mechanizmov. Väčšina štúdií skúmala najmä postherpetickú neuralgiu a bolestivé diabetické neuropatie, ale výsledky nemusia platiť pre všetky stavy neuropatickej bolesti.
Antidepresíva
Antidepresíva zvyšujú hladiny synaptického serotonínu a norepinefrínu, čím zosilňujú účinok zostupného analgetického systému spojeného s neuropatickou bolesťou. Boli základom terapie neuropatickej bolesti. Analgetické účinky možno pripísať blokáde spätného vychytávania nor-adrenalínu a dopamínu, ktorá pravdepodobne zvyšuje zostupnú inhibíciu, antagonizmus NMDA-receptorov a blokádu sodíkových kanálov. tricyklické antidepresíva, ako sú TCA; napr. amitriptylín, imipramín, nortriptylín a doxepín sú účinné proti neustálej bolestivej alebo pálivej bolesti spolu so spontánnou bolesťou.
Ukázalo sa, že tricyklické antidepresíva sú výrazne účinnejšie pri neuropatickej bolesti ako špecifické inhibítory spätného vychytávania serotonínu alebo SSRI, ako je fluoxetín, paroxetín, sertralín a citalopram. Dôvodom môže byť, že inhibujú spätné vychytávanie serotonínu a norepinefrínu, zatiaľ čo SSRI inhibujú iba spätné vychytávanie serotonínu. Tricyklické antidepresíva môžu mať nepríjemné vedľajšie účinky, vrátane nevoľnosti, zmätenosti, blokády srdcového vedenia, tachykardie a ventrikulárnych arytmií. Môžu tiež spôsobiť zvýšenie telesnej hmotnosti, znížený prah záchvatov a ortostatickú hypotenziu. Tricyklické lieky sa musia používať opatrne u starších ľudí, ktorí sú obzvlášť náchylní na ich akútne vedľajšie účinky. Koncentrácia liečiva v krvi sa má monitorovať, aby sa predišlo toxicite u pacientov, ktorí sú pomalými metabolizátormi liekov.
Inhibítory spätného vychytávania serotonínu a norepinefrínu alebo SNRI sú novou triedou antidepresív. Rovnako ako TCA sa zdajú byť účinnejšie ako SSRI na liečbu neuropatickej bolesti, pretože tiež inhibujú spätné vychytávanie norepinefrínu a dopamínu. Venlafaxín je rovnako účinný proti oslabujúcim polyneuropatiám, ako je bolestivá diabetická neuropatia, ako imipramín, pri zmienke o TCA, a tieto dva účinky sú výrazne vyššie ako placebo. Zdá sa, že podobne ako TCA, aj SNRI poskytujú výhody nezávislé od ich antidepresívnych účinkov. Vedľajšie účinky zahŕňajú sedáciu, zmätenosť, hypertenziu a abstinenčný syndróm.
Antiepileptické lieky
Antiepileptiká môžu byť použité ako liečba prvej línie najmä pri niektorých typoch neuropatickej bolesti. Pôsobia tak, že modulujú napäťovo riadené vápnikové a sodíkové kanály, zlepšujú inhibičné účinky GABA a inhibujú excitačný glutamínergný prenos. Antiepileptiká sa nepreukázali ako účinné pri akútnej bolesti. V prípadoch chronickej bolesti sa zdá, že antiepileptiká sú účinné iba pri neuralgii trojklanného nervu. Na tento stav sa rutinne používa karbamazepín. Gabapentín, ktorý pôsobí inhibíciou funkcie vápnikového kanála prostredníctvom agonistických účinkov na alfa-2 delta podjednotke vápnikového kanála, je tiež známy ako účinný pri neuropatickej bolesti. Gabapentín však pôsobí centrálne a môže spôsobiť únavu, zmätenosť a somnolenciu.
Neopioidné analgetiká
Chýbajú silné údaje podporujúce používanie nesteroidných protizápalových liekov alebo NSAID pri úľave od neuropatickej bolesti. Môže to byť spôsobené nedostatkom zápalovej zložky pri zmierňovaní bolesti. Používali sa však zameniteľne s opioidmi ako adjuvans pri liečbe bolesti pri rakovine. Boli však hlásené komplikácie, najmä u ťažko oslabených pacientov.
Opioidné analgetiká
Opioidné analgetiká sú predmetom mnohých diskusií pri zmierňovaní neuropatickej bolesti. Pôsobia inhibíciou centrálnych vzostupných bolestivých impulzov. Tradične sa predtým pozorovalo, že neuropatická bolesť je odolná voči opioidom, pričom opioidy sú vhodnejšími metódami pre koronárne a somatické nociceptívne typy bolesti. Mnoho lekárov bráni používaniu opioidov na liečbu neuropatickej bolesti, z veľkej časti kvôli obavám zo zneužívania drog, závislosti a regulačných problémov. Existuje však mnoho štúdií, ktoré ukázali, že opioidné analgetiká sú úspešné. Oxykodón bol lepší ako placebo na zmiernenie bolesti, alodýnie, zlepšenie spánku a hendikepu. Opioidy s riadeným uvoľňovaním podľa plánu sa odporúčajú pacientom s neustálou bolesťou, aby podporili konštantné hladiny analgézie, zabránili kolísaniu hladiny glukózy v krvi a zabránili nežiaducim udalostiam spojeným s vyšším dávkovaním. Najčastejšie sa perorálne prípravky používajú kvôli ich ľahšiemu použitiu a nákladovej efektívnosti. Transdermálne, parenterálne a rektálne prípravky sa vo všeobecnosti používajú u pacientov, ktorí netolerujú perorálne lieky.
Lokálne anestetiká
Neďaleko pôsobiace anestetiká sú príťažlivé, pretože vďaka svojmu regionálnemu účinku majú minimálne vedľajšie účinky. Pôsobia stabilizáciou sodíkových kanálov na axónoch periférnych neurónov prvého poriadku. Fungujú najlepšie, ak dôjde len k čiastočnému poškodeniu nervov a nahromadia sa prebytočné sodíkové kanály. Topický lidokaín je najlepšie študovaným predstaviteľom kurzu pre neuropatickú bolesť. Konkrétne použitie tejto 5-percentnej lidokaínovej náplasti na postherpetickú neuralgiu spôsobilo jej schválenie FDA. Zdá sa, že náplasť funguje najlepšie, keď je poškodená, ale udržiavaná funkcia nociceptora periférneho nervového systému z príslušného dermatómu, ktorá sa prejavuje ako alodýnia. Je potrebné ho aplikovať priamo na symptomatickú oblasť na 12 hodín a odstraňovať ďalších 12 hodín a takto sa môže používať roky. Okrem lokálnych kožných reakcií je často dobre tolerovaný mnohými pacientmi s neuropatickou bolesťou.
Rôzne Drogy
Ukázalo sa, že klonidín, alfa-2-agonista, je účinný u podskupiny pacientov s diabetickou periférnou neuropatiou. Zistilo sa, že kanabinoidy zohrávajú úlohu v experimentálnej modulácii bolesti na zvieracích modeloch a dôkazy o účinnosti sa hromadia. CB2-selektívne agonisty potláčajú hyperalgéziu a alodýniu a normalizujú nociceptívne prahy bez vyvolania analgézie.
Intervenčný manažment bolesti
U pacientov, ktorí majú nezvládnuteľnú neuropatickú bolesť, možno zvážiť invazívnu liečbu. Tieto liečby zahŕňajú epidurálne alebo perineurálne injekcie lokálnych anestetík alebo kortikosteroidov, implantáciu epidurálnych a intratekálnych metód podávania liekov a zavedenie stimulátorov miechy. Tieto prístupy sú vyhradené pre pacientov s nezvládnuteľnou chronickou neuropatickou bolesťou, u ktorých zlyhala konzervatívna liečba a tiež prešli dôkladným psychologickým vyšetrením. V štúdii Kim et al sa ukázalo, že stimulátor miechy bol účinný pri liečbe neuropatickej bolesti pôvodom z nervových koreňov.
Pohľad Dr. Alexa Jimeneza
Pri neuropatickej bolesti sa symptómy chronickej bolesti vyskytujú v dôsledku poškodenia, dysfunkčnosti alebo poranenia samotných nervových vlákien, čo je vo všeobecnosti sprevádzané poškodením alebo poranením tkaniva. Výsledkom je, že tieto nervové vlákna môžu začať vysielať nesprávne signály bolesti do iných oblastí tela. Účinky neuropatickej bolesti spôsobenej poranením nervových vlákien zahŕňajú modifikácie funkcie nervov v mieste poranenia aj v oblastiach okolo poranenia. Pochopenie patofyziológie neuropatickej bolesti bolo cieľom mnohých zdravotníckych pracovníkov, aby mohli efektívne určiť najlepší liečebný prístup, ktorý pomôže zvládnuť a zlepšiť jej symptómy. Od užívania liekov a/alebo liekov až po chiropraktickú starostlivosť, cvičenie, fyzickú aktivitu a výživu možno použiť rôzne liečebné prístupy, ktoré pomôžu zmierniť neuropatickú bolesť pre potreby každého jednotlivca.
Ďalšie intervencie pre neuropatickú bolesť
Mnoho pacientov s neuropatickou bolesťou využíva doplnkové a alternatívne možnosti liečby na liečbu neuropatickej bolesti. Iné dobre známe režimy používané na liečbu neuropatickej bolesti zahŕňajú akupunktúru, perkutánnu elektrickú nervovú stimuláciu, transkutánnu elektrickú nervovú stimuláciu, kognitívno-behaviorálnu liečbu, odstupňované motorické zobrazovanie a podpornú liečbu a cvičenie. Medzi nimi je však chiropraktická starostlivosť dobre známym alternatívnym liečebným prístupom, ktorý sa bežne používa na liečbu neuropatickej bolesti. Chiropraktická starostlivosť spolu s fyzikálnou terapiou, cvičením, úpravou výživy a životného štýlu môže v konečnom dôsledku ponúknuť úľavu od symptómov neuropatickej bolesti.
Chiropraxe starostlivosti
Je známe, že komplexná aplikácia manažmentu je rozhodujúca v boji proti účinkom neuropatickej bolesti. Týmto spôsobom je chiropraktická starostlivosť holistický liečebný program, ktorý by mohol byť účinný pri prevencii zdravotných problémov spojených s poškodením nervov. Chiropraktická starostlivosť poskytuje pomoc pacientom s mnohými rôznymi stavmi, vrátane pacientov s neuropatickou bolesťou. Pacienti trpiaci neuropatickou bolesťou často používajú nesteroidné protizápalové lieky alebo NSAID, ako je ibuprofén, alebo ťažké lieky proti bolesti na predpis, aby pomohli zmierniť neuropatickú bolesť. Môžu poskytnúť dočasnú opravu, ale na zvládnutie bolesti je potrebné ich neustále používať. To vždy prispieva k škodlivým vedľajším účinkom av extrémnych situáciách k závislosti od liekov na predpis.
Chiropraktická starostlivosť môže pomôcť zlepšiť symptómy neuropatickej bolesti a zvýšiť stabilitu bez týchto nevýhod. Prístup, akým je chiropraktická starostlivosť, ponúka individuálny program navrhnutý tak, aby presne určil hlavnú príčinu problému. Pomocou úprav chrbtice a manuálnych manipulácií môže chiropraktik starostlivo napraviť akékoľvek vychýlenie chrbtice alebo subluxácie, ktoré sa nachádzajú po celej dĺžke chrbtice, čo by mohlo znížiť následky poškodenia nervov prostredníctvom prestavby chrbtice. Obnovenie integrity chrbtice je nevyhnutné na udržanie vysoko funkčného centrálneho nervového systému.
Chiropraktik môže byť tiež dlhodobá liečba zameraná na zlepšenie vašej celkovej pohody. Okrem úprav chrbtice a manuálnych manipulácií môže chiropraktik ponúknuť výživové poradenstvo, ako je napríklad predpisovanie stravy bohatej na antioxidanty, alebo môže navrhnúť fyzikálnu terapiu alebo cvičebný program na boj proti vzplanutiu bolesti nervov. Dlhodobý stav si vyžaduje dlhodobú nápravu a v tejto funkcii môže byť zdravotnícky pracovník, ktorý sa špecializuje na zranenia a/alebo stavy ovplyvňujúce pohybový a nervový systém, ako napríklad lekár chiropraktika alebo chiropraktik, pri svojej práci neoceniteľný. na meranie priaznivých zmien v priebehu času.
Fyzikálna terapia, cvičenie a techniky reprezentácie pohybu sa ukázali ako prospešné pri liečbe neuropatickej bolesti. Chiropraktická starostlivosť ponúka aj iné liečebné modality, ktoré môžu byť užitočné pri zvládaní alebo zlepšovaní neuropatickej bolesti. Nízkoúrovňová laserová terapia alebo LLLT, napríklad, získala obrovský význam ako liečba neuropatickej bolesti. Podľa rôznych výskumných štúdií sa dospelo k záveru, že LLLT má pozitívne účinky na kontrolu analgézie pri neuropatickej bolesti, sú však potrebné ďalšie výskumné štúdie na definovanie liečebných protokolov, ktoré sumarizujú účinky nízkoúrovňovej laserovej terapie pri liečbe neuropatickej bolesti.
Chiropraktická starostlivosť zahŕňa aj výživové poradenstvo, ktoré môže pomôcť kontrolovať symptómy spojené s diabetickou neuropatiou. Počas výskumnej štúdie sa preukázalo, že nízkotučná rastlinná strava zlepšuje kontrolu glykémie u pacientov s cukrovkou 2. typu. Po približne 20 týždňoch pilotnej štúdie jednotlivci, ktorých sa to týka, hlásili zmeny telesnej hmotnosti a elektrochemická vodivosť kože na chodidle sa po intervencii zlepšila. Výskumná štúdia naznačila potenciálnu hodnotu intervencie nízkotučnej rastlinnej stravy pri diabetickej neuropatii. Okrem toho klinické štúdie zistili, že perorálna aplikácia magnézium L-treonátu je schopná zabrániť, ako aj obnoviť pamäťové deficity spojené s neuropatickou bolesťou.
Chiropraktická starostlivosť môže tiež ponúknuť ďalšie liečebné stratégie na podporu regenerácie nervov. Napríklad bolo navrhnuté, že zvýšenie regenerácie axónov pomôže zlepšiť funkčné zotavenie po poranení periférnych nervov. Podľa nedávnych výskumných štúdií sa zistilo, že elektrická stimulácia spolu s cvičením alebo fyzickými aktivitami podporuje regeneráciu nervov po oneskorenej oprave nervov u ľudí a potkanov. Elektrická stimulácia aj cvičenie boli nakoniec určené ako sľubné experimentálne liečby poškodenia periférnych nervov, ktoré sa zdajú byť pripravené na klinické použitie. Na úplné určenie ich účinkov u pacientov s neuropatickou bolesťou môžu byť potrebné ďalšie výskumné štúdie.
záver
Neuropatická bolesť je mnohostranná entita bez osobitných usmernení, o ktoré sa treba starať. Najlepšie sa to dá zvládnuť pomocou multidisciplinárneho prístupu. Manažment bolesti si vyžaduje neustále hodnotenie, edukáciu pacienta, zabezpečenie sledovania pacienta a uistenie. Neuropatická bolesť je chronický stav, ktorý sťažuje možnosť najlepšej liečby. Individualizácia liečby zahŕňa posúdenie vplyvu bolesti na pohodu, depresiu a postihnutie jednotlivca spolu s ďalším vzdelávaním a hodnotením. Štúdie neuropatickej bolesti, ako na molekulárnej úrovni a na zvieracích modeloch, je relatívne nová, ale veľmi sľubná. V základných a klinických oblastiach neuropatickej bolesti sa očakávajú mnohé zlepšenia, čím sa otvárajú dvere k zlepšeným alebo novým liečebným modalitám pre tento invalidizujúci stav. Rozsah našich informácií je obmedzený na chiropraktiku, ako aj na poranenia a stavy chrbtice. Ak chcete prediskutovať túto tému, neváhajte sa opýtať Dr. Jimeneza alebo nás kontaktujte na adrese�915-850-0900 .
Kurátorom je Dr. Alex Jimenez
Ďalšie témy: Bolesť chrbta
Bolesti chrbta je jednou z najčastejších príčin invalidity a vymeškaných dní v práci na celom svete. Bolesti chrbta sa v skutočnosti pripisujú ako druhý najčastejší dôvod návštevy lekára, prevyšujú ju len infekcie horných dýchacích ciest. Približne 80 percent populácie aspoň raz za život zažije nejaký typ bolesti chrbta. Chrbtica je zložitá štruktúra pozostávajúca z kostí, kĺbov, väzov a svalov, okrem iného z mäkkých tkanív. Z tohto dôvodu môžu zranenia a/alebo zhoršené stavy, ako napr herné disky, môže nakoniec viesť k príznakom bolesti chrbta. Športové zranenia alebo zranenia pri automobilových nehodách sú často najčastejšou príčinou bolesti chrbta, niekedy však môžu mať aj tie najjednoduchšie pohyby bolestivé následky. Našťastie alternatívne možnosti liečby, ako je chiropraktická starostlivosť, môžu pomôcť zmierniť bolesť chrbta pomocou úprav chrbtice a manuálnych manipulácií, čo v konečnom dôsledku zlepšuje úľavu od bolesti.
Bolesť Úzkosť Depresia�Každý zažil bolesť, existujú však ľudia s depresiou, úzkosťou alebo oboje. Skombinujte to s bolesťou a môže byť dosť intenzívna a ťažko liečiteľná. Ľudia, ktorí trpia depresiou, úzkosťou alebo oboma, majú tendenciu pociťovať silnú a dlhodobú bolesť viac ako iní ľudia.
Cesta úzkosť, Depresia a bolesť sa navzájom prekrývajú v chronickej a v niektorých syndrómoch bolesti, tj nízka bolesť chrbta, bolesti hlavybolesť nervov a fibromyalgia. Psychiatrické poruchy prispievajú k intenzite bolesti a tiež zvyšujú riziko invalidity.
depresia:�A (veľká depresívna porucha alebo klinická depresia) je bežná, ale závažná porucha nálady. Spôsobuje vážne symptómy, ktoré ovplyvňujú to, ako sa jednotlivec cíti, myslí a ako zvláda každodenné činnosti, tj spánok, jedenie a prácu. Aby bola diagnostikovaná depresia, symptómy musia byť prítomné aspoň dva týždne.
Pretrvávajúca smutná, úzkostná alebo „prázdna“ nálada.
Pocit beznádeje, pesimizmus.
Podráždenosť.
Pocit viny, bezcennosti alebo bezmocnosti.
Strata záujmu alebo potešenia z aktivít.
Znížená energia alebo únava.
Pohybujte sa alebo rozprávajte pomaly.
Pocit nepokoja a problémy s pokojným sedením.
Ťažkosti so sústredením, zapamätaním alebo rozhodovaním.
Ťažkosti so spánkom, ranné prebúdzanie a zaspávanie.
Zmeny chuti do jedla a hmotnosti.
Myšlienky na smrť alebo samovraždu alebo pokusy o samovraždu.
Bolesti alebo bolesti, bolesti hlavy, kŕče alebo tráviace problémy bez jasnej fyzickej príčiny a/alebo ktoré neuľahčujú liečbou.
Nie každý, kto má depresiu, zažíva všetky príznaky. Niektorí pociťujú len niekoľko príznakov, zatiaľ čo iní môžu mať niekoľko. Okrem nízkej nálady je niekoľko pretrvávajúcich symptómov�potrebný�pre diagnózu ťažkej depresie. Závažnosť a frekvencia symptómov spolu s trvaním sa budú líšiť v závislosti od jednotlivca a jeho konkrétneho ochorenia. Príznaky sa môžu líšiť aj v závislosti od štádia ochorenia.
BOLESŤ ÚZKOSŤ DEPRESIA
ciele:
Aký je vzťah?
Aká je neurofyziológia za tým?
Aké sú hlavné dôsledky?
Zmeny v mozgu pri bolesti
Obrázok 1 Mozgové dráhy, oblasti a siete zapojené do akútnej a chronickej bolesti
Davis, KD a kol. (2017) Zobrazovacie testy mozgu na chronickú bolesť: lekárske, právne a etické problémy a odporúčania Nat. Neurol. doi:10.1038/nrneurol.2017.122
BOLESŤ, ÚZKOST A DEPRESIA
záver:
Bolesť, najmä chronická, je spojená s depresiou a úzkosťou
Fyziologické mechanizmy vedúce k úzkosti a depresii môžu mať multifaktoriálny charakter
Keď je zmyslový systém ovplyvnený zranením alebo chorobou, nervy v tomto systéme nemôžu správne fungovať, aby prenášali pocity a pocity do mozgu. To často prispieva k pocitu necitlivosti alebo nedostatku citlivosti. Avšak, v niektorých prípadoch, keď je tento systém poškodený, ľudia môžu pociťovať bolesť v postihnutej oblasti.
Neuropatická bolesť nezačína náhle ani sa rýchlo nevyrieši; je to a chronická bolesť stav, ktorý vedie k symptómom pretrvávajúcej bolesti. U väčšiny jedincov sa intenzita symptómov môže stupňovať a slabnúť počas dňa. Hoci sa predpokladá, že neuropatická bolesť súvisí so zdravotnými problémami periférnych nervov, ako je neuropatia spôsobená cukrovkou alebo spinálnou stenózou, poranenia mozgu alebo miechy môžu tiež viesť k chronickej neuropatickej bolesti. Neuropatická bolesť sa označuje aj ako bolesť nervov.
Neuropatická bolesť môže byť v kontraste s nociceptívnou bolesťou. Neuropatická bolesť sa nevyvinie na základe žiadnych špecifických okolností alebo vonkajšieho podnetu, ale symptómy sa vyskytujú jednoducho preto, že nervový systém nemusí fungovať zodpovedajúcim spôsobom. V skutočnosti môžu jednotlivci pociťovať neuropatickú bolesť aj vtedy, keď sa boľavá alebo poranená časť tela v skutočnosti nenachádza. Tento stav sa nazýva fantómová bolesť končatín, ktorá sa môže vyskytnúť u ľudí po amputácii.
Nociceptívna bolesť je vo všeobecnosti akútna a vyvíja sa v reakcii na špecifickú okolnosť, napríklad keď niekto utrpí náhle zranenie, ako je udieranie do prsta kladivom alebo narážanie palcom pri chôdzi naboso. Okrem toho nociceptívna bolesť má tendenciu ustúpiť, keď sa postihnuté miesto zahojí. Telo obsahuje špecializované nervové bunky, známe ako nociceptory, ktoré detegujú škodlivé podnety, ktoré by mohli poškodiť telo, ako je extrémne teplo alebo chlad, tlak, zvieranie a vystavenie chemikáliám. Tieto varovné signály sa potom prenášajú pozdĺž nervového systému do mozgu, čo vedie k nocicepčnej bolesti.
Aké sú rizikové faktory neuropatickej bolesti?
Čokoľvek, čo prispieva k nedostatočnej funkcii zmyslového nervového systému, môže viesť k neuropatickej bolesti. Problémy so zdravím nervov zo syndrómu karpálneho tunela alebo podobných stavov môžu nakoniec vyvolať neuropatickú bolesť. Trauma, ktorá vedie k poškodeniu nervov, môže viesť k neuropatickej bolesti. Ďalšie stavy, ktoré môžu predisponovať jednotlivcov k rozvoju neuropatickej bolesti, zahŕňajú: cukrovku, nedostatok vitamínov, rakovinu, HIV, mŕtvicu, roztrúsenú sklerózu, pásový opar a dokonca aj niektoré liečby rakoviny.
Aké sú príčiny neuropatickej bolesti?
Existuje mnoho príčin, z ktorých sa u jednotlivcov môže vyvinúť neuropatická bolesť. Ale na bunkovej úrovni je jedným z vysvetlení zvýšené uvoľňovanie určitých receptorov, ktoré indikujú bolesť, spolu so zníženou schopnosťou nervov modulovať tieto signály, čo vedie k pocitu bolesti pochádzajúcej z postihnutej oblasti. Okrem toho sa v mieche preskupuje oblasť, ktorá má bolestivé príznaky, so zodpovedajúcimi zmenami hormónov a stratou normálne fungujúcich pohyblivých tiel. Tieto zmeny vedú k vnímaniu bolesti pri absencii vonkajšej stimulácie. V mozgu, schopnosť blokovať bolesť môže byť ovplyvnená po zranení, ako je mŕtvica alebo trauma z poranenia. Postupom času dochádza k ďalšiemu poškodeniu buniek a pocit bolesti pokračuje. Neuropatická bolesť súvisí aj s cukrovkou, chronickým príjmom alkoholu, niektorými druhmi rakoviny, nedostatkom vitamínu B, chorobami, inými nervovými chorobami, toxínmi a špecifickými liekmi.
Aké sú príznaky neuropatickej bolesti?
Na rozdiel od iných neurologických stavov môže byť identifikácia neuropatickej bolesti náročná. Môže však byť prítomných niekoľko objektívnych signálov, ak vôbec nejaké. Zdravotnícki pracovníci musia rozlúštiť a preložiť celý rad slov, ktoré pacienti používajú na opis svojej bolesti. Pacienti môžu opísať svoje symptómy ako ostré, tupé, horúce, studené, citlivé, svrbiace, hlboké, štípanie, pálenie, medzi rôznymi inými popisnými termínmi. Niektorí pacienti môžu navyše pociťovať bolesť ľahkým dotykom alebo tlakom.
V snahe pomôcť identifikovať, akú veľkú bolesť by pacienti mohli podstúpiť, sa často používajú rôzne škály. Pacienti sú požiadaní, aby ohodnotili svoju bolesť podľa vizuálnej stupnice alebo číselného grafu. Existuje mnoho príkladov stupníc bolesti, ako je ten, ktorý je uvedený nižšie. Obrázky tvárí zobrazujúce rôzne úrovne bolesti môžu byť často užitočné, keď majú jednotlivci problém opísať množstvo bolesti, ktorú zažívajú.
Chronická bolesť a duševné zdravie
Pre mnohých nemusí byť vplyv chronickej bolesti obmedzený na bolesť; môže to negatívne ovplyvniť aj ich psychický stav. Nové výskumné štúdie vedcov z Northwestern University v Chicagu môžu vysvetliť, prečo jednotlivci, ktorí majú chronickú bolesť, trpia aj zdanlivo nesúvisiacimi zdravotnými problémami, ako sú depresia, stres, nedostatok spánku a problémy so sústredením.
Hodnotenie ukázalo, že ľudia s chronickou bolesťou vykazujú rôzne oblasti mozgu, ktoré sú vždy aktívne, najmä oblasť spojenú s náladou a pozornosťou. Toto nepretržité pôsobenie prepája nervové spojenia z mozgu a necháva ľudí trpiacich chronickou bolesťou vo vyššie riziko psychických problémov. Výskumníci naznačili, že neustále prijímanie signálov bolesti môže viesť k mentálnemu prepojeniu, ktoré nepriaznivo ovplyvňuje myseľ. Preinštalovanie núti ich mozgy, aby venovali svoje mentálne zdroje iným spôsobom na riešenie každodenných úloh, od matematiky, cez zapamätanie si nákupného zoznamu až po pocit šťastia.
Spojenie medzi bolesťou a mozgom bolo dobre zaznamenané, prinajmenšom neoficiálne, a mnohí zdravotnícki pracovníci tvrdia, že z prvej ruky videli, ako môže duševný stav pacienta ísť z kopca, keď znáša chronickú bolesť. Mylné predstavy o prepojení bolesť-mozog mohli vyplynúť z nedostatku dôkazov, že bolesť má merateľný a trvalý vplyv na mozog. Výskumníci očakávajú, že s ďalším výskumom mechanizmov toho, ako chronická bolesť spôsobuje, že ľudia sú náchylnejší na poruchy nálady, budú mať ľudia schopnosť lepšie zvládať svoju celkovú pohodu.
Kultúra a chronická bolesť
Veľa vecí prispieva k tomu, ako prežívame a vyjadrujeme bolesť, no nedávno výskumníci naznačili, že kultúra priamo súvisí s prejavom bolesti. Naše výchovné a spoločenské hodnoty ovplyvňujú, ako vyjadrujeme bolesť a tiež vlastnú povahu, intenzitu a dĺžku. Tieto premenné však nie sú také zrejmé ako sociálno-psychologické hodnoty, ako je vek a pohlavie.
Výskum uvádza, že chronická bolesť je mnohostranný proces a súbežná súhra medzi patofyziológiou, kognitívnymi, afektívnymi, behaviorálnymi a sociokultúrnymi faktormi sa spája s tým, čo sa označuje ako skúsenosť s chronickou bolesťou. Ukázalo sa, že chronická bolesť sa u pacientov rôznych kultúr a etník prejavuje odlišne.
Niektoré kultúry podporujú prejav bolesti, najmä v južnom Stredomorí a na Strednom východe. Iní jednotlivci to potláčajú, ako v mnohých lekciách pre naše deti o statočnom správaní a neplakaní. Bolesť je uznávaná ako súčasť ľudskej skúsenosti. Sme náchylní predpokladať, že komunikácia o bolesti bez problémov prekročí kultúrne hranice. Ľudia trpiaci bolesťou však podliehajú spôsobom, ktoré ich civilizácia vycvičila, aby prežívali a vyjadrovali bolesť.
Jedinci s bolesťou aj zdravotnícki pracovníci majú ťažkosti s komunikáciou bolesti cez etnické hranice. Vo veciach, ako je bolesť, kde efektívna komunikácia môže mať ďalekosiahle dôsledky pre lekársku starostlivosť, kvalitu života a potenciálne prežitie, zostáva úloha kultúry pri komunikácii bolesti podhodnotená. Pretrvávajúca bolesť je multidimenzionálnym, zloženým stretnutím tvoreným prelínaním a spolupôsobením biologických a psychosociálnych faktorov. Poznanie kulminácie týchto faktorov je rozhodujúce pre pochopenie rozdielov v ich prejavoch a riadení.
Ako sa diagnostikuje neuropatická bolesť?
Diagnóza neuropatickej bolesti sa spolieha na dodatočné hodnotenie histórie jednotlivca. Ak je podozrenie na podozrenie na podozrenie, potom sa môže odôvodniť analýza nervov spolu s testovaním. Najbežnejším prostriedkom na posúdenie, či je alebo nie je poškodený nerv, používa elektrodiagnostický liek. Táto lekárska subspecialita využíva techniky štúdií nervov s elektromyelografiou (Ncs / EMG). Klinické hodnotenie môže preukázať stratu práce a môže zahŕňať hodnotenie ľahkého dotyku, schopnosti rozlíšiť ostrú bolesť od tupej bolesti a schopnosti rozlíšiť teplotu, ako aj hodnotenie vibrácií.
Po dôkladnom klinickom vyšetrení je možné naplánovať elektrodiagnostickú analýzu. Tieto štúdie vykonávajú špeciálne vyškolení neurológovia a fyziatri. Ak existuje podozrenie na neuropatiu, malo by sa začať hľadať reverzibilné príčiny. To môže zahŕňať funkciu krvi pre nedostatok vitamínov alebo problémy so štítnou žľazou a zobrazovacie štúdie na vylúčenie štrukturálnej lézie postihujúcej miechu. V závislosti od výsledkov tohto testovania môžu existovať prostriedky na zníženie intenzity neuropatie a možno aj na zníženie bolesti, ktorú pacient podstupuje.
Žiaľ, v mnohých stavoch ani dobrá kontrola základnej príčiny neuropatie nemôže zvrátiť neuropatickú bolesť. Toto sa bežne pozoruje u pacientov s diabetickou neuropatiou. V zriedkavých prípadoch sa môžu v postihnutej oblasti vyskytnúť známky zmien v raste kože a ochlpenia. Tieto zmeny môžu byť spojené so zmenami v potení. Ak sú prítomné, tieto zmeny môžu pomôcť identifikovať pravdepodobnú prítomnosť neuropatickej bolesti súvisiacej so stavom známym ako syndróm komplexnej regionálnej bolesti.
Pohľad Dr. Alexa Jimeneza
Neuropatická bolesť je chronický bolestivý stav, ktorý je vo všeobecnosti spojený s priamym poškodením alebo poranením nervového systému alebo nervov. Tento typ bolesti sa líši od nociceptívnej bolesti alebo typického pocitu bolesti. Nociceptívna bolesť je akútny alebo náhly pocit bolesti, ktorý spôsobuje, že nervový systém vysiela signály bolesti ihneď po tom, ako došlo k traume. Pri neuropatickej bolesti však môžu pacienti pociťovať vystreľujúcu, pálivú bolesť bez akéhokoľvek priameho poškodenia alebo poranenia. Pochopenie možných príčin pacientovej neuropatickej bolesti v porovnaní s akýmkoľvek iným typom bolesti môže zdravotníckym pracovníkom pomôcť nájsť lepšie spôsoby liečby chronických bolestivých stavov.
Aká je liečba neuropatickej bolesti?
Na liečbu neuropatickej bolesti sa používajú rôzne lieky. Väčšina týchto liekov sa používa mimo označenia, čo znamená, že liek bol schválený FDA na liečbu rôznych stavov a potom bol uznaný ako výhodný na liečbu neuropatickej bolesti. Tricyklické antidepresíva, ako je amitriptylín, nortriptylín a desipramín, sa už niekoľko rokov predpisujú na liečbu neuropatickej bolesti.
Niektorí jednotlivci zistia, že tieto môžu byť veľmi účinné pri poskytovaní úľavy. Ukázalo sa, že iné druhy antidepresív ponúkajú určitú úľavu. U niektorých pacientov sa používajú selektívne inhibítory spätného vychytávania serotonínu alebo SSRI, ako je paroxetín a citalopram, a iné antidepresíva, ako je venlafaxín a bupropión. Ďalšia častá liečba neuropatickej bolesti zahŕňa lieky proti záchvatom, vrátane karbamazepínu, fenytoínu, gabapentínu, lamotrigínu a ďalších.
V akútnych prípadoch bolestivej neuropatie, ktorá nereaguje na sprostredkovateľov prvej línie, môžu byť lieky zvyčajne používané na liečbu srdcových arytmií určitým prínosom; tieto však môžu viesť k významným vedľajším účinkom a často sa musia pozorne sledovať. Lieky aplikované priamo na kožu môžu niektorým pacientom ponúknuť mierny až citeľný prínos. Bežne používané formy zahŕňajú lidokaín (vo forme náplasti alebo gélu) alebo kapsaicín.
Liečba neuropatickej bolesti závisí od základnej príčiny. Ak je príčina reverzibilná, potom sa periférne nervy môžu regenerovať a bolesť ustúpi; toto zníženie bolesti však môže trvať niekoľko mesiacov až rokov. Môže sa tiež použiť niekoľko ďalších alternatívnych možností liečby, vrátane chiropraktickej starostlivosti a fyzikálnej terapie, aby sa pomohlo zmierniť napätie a stres pozdĺž nervov, čo v konečnom dôsledku pomôže zlepšiť bolestivé symptómy.
Aká je prognóza neuropatickej bolesti?
Mnohí jedinci s neuropatickou bolesťou sú schopné získať určitú mieru pomoci, aj keď ich bolesť pretrváva. Hoci neuropatická bolesť nie je pre pacienta nebezpečná, prítomnosť chronickej bolesti môže negatívne ovplyvniť kvalitu života. Pacienti s chronickou bolesťou nervov môžu trpieť nedostatkom spánku alebo poruchami nálady, vrátane depresie, úzkosti a stresu, ako už bolo uvedené vyššie. Vzhľadom na inherentnú alopéciu a nedostatok senzorickej spätnej väzby sú pacienti vystavení riziku vzniku zranenia alebo infekcie alebo nevedomky spôsobia eskaláciu súčasného zranenia. Preto je nevyhnutné okamžite vyhľadať lekársku pomoc a dodržiavať špecifické pokyny zdravotníckeho pracovníka pre bezpečnosť a opatrnosť.
Môže byť zabránené neuropatickej bolesti?
Najlepším spôsobom, ako zabrániť neuropatickej bolesti, je vyhnúť sa rozvoju alebo progresii neuropatie. Monitorovanie a zmena možností životného štýlu vrátane obmedzenia konzumácie alkoholu a tabaku; udržiavanie zdravej hmotnosti, aby sa znížilo riziko cukrovky, degeneratívneho ochorenia kĺbov alebo mŕtvice; a mať skvelú ergonomickú formu v práci alebo pri vykonávaní koníčkov na zníženie rizika opakovaného stresového zranenia sú stratégie na zníženie pravdepodobnosti rozvoja neuropatie a potenciálnej neuropatickej bolesti. V prípade akýchkoľvek symptómov spojených s neuropatickou bolesťou okamžite vyhľadajte lekársku pomoc, aby ste mohli zvoliť najvhodnejší liečebný prístup.�Rozsah našich informácií je obmedzený na chiropraxiu, ako aj na poranenia a stavy chrbtice. Ak chcete prediskutovať túto tému, neváhajte sa opýtať Dr. Jimeneza alebo nás kontaktujte na adrese�915-850-0900 .
Kurátorom je Dr. Alex Jimenez
Ďalšie témy: Bolesť chrbta
Bolesti chrbta je jednou z najčastejších príčin invalidity a vymeškaných dní v práci na celom svete. Bolesti chrbta sa v skutočnosti pripisujú ako druhý najčastejší dôvod návštevy lekára, prevyšujú ju len infekcie horných dýchacích ciest. Približne 80 percent populácie aspoň raz za život zažije nejaký typ bolesti chrbta. Chrbtica je zložitá štruktúra pozostávajúca z kostí, kĺbov, väzov a svalov, okrem iného z mäkkých tkanív. Z tohto dôvodu môžu zranenia a/alebo zhoršené stavy, ako napr herné disky, môže nakoniec viesť k príznakom bolesti chrbta. Športové zranenia alebo zranenia pri automobilových nehodách sú často najčastejšou príčinou bolesti chrbta, niekedy však môžu mať aj tie najjednoduchšie pohyby bolestivé následky. Našťastie alternatívne možnosti liečby, ako je chiropraktická starostlivosť, môžu pomôcť zmierniť bolesť chrbta pomocou úprav chrbtice a manuálnych manipulácií, čo v konečnom dôsledku zlepšuje úľavu od bolesti.
Ak je zmyslový systém ovplyvnený zranením alebo chorobou, nervy v tomto systéme nemôžu fungovať pri prenose pocitov do mozgu. To môže viesť k pocitu necitlivosti alebo nedostatku citlivosti. V niektorých prípadoch, keď je zmyslový systém poškodený, môžu jednotlivci pociťovať bolesť v postihnutej oblasti. Neuropatická bolesť nezačína rýchlo alebo rýchlo končí. Ide o chronický stav, ktorý vedie k príznakom pretrvávajúca bolesť. Pre mnohých môže intenzita symptómov prísť a odísť počas dňa. Predpokladá sa, že neuropatická bolesť súvisí s problémami periférnych nervov, tj neuropatia spôsobená cukrovkou, spinálna stenóza, poranenie mozgu alebo miechy môže tiež viesť k chronickej neuropatickej bolesti.
NEUROPATICKÁ BOLESŤ
ciele:
Čo je to?
Aká je za tým patofyziológia?
Aké sú príčiny
Aké sú niektoré z ciest
Ako to môžeme opraviť?
NEUROPATICKÁ BOLESŤ
Bolesť iniciovaná alebo spôsobená primárnou léziou alebo dysfunkciou v somatosenzorickom nervovom systéme.
Neuropatická bolesť je zvyčajne chronická, ťažko liečiteľná a často odolná voči štandardnej analgetickej liečbe.
PATOGENÉZA NEUROPATICKEJ BOLESTI
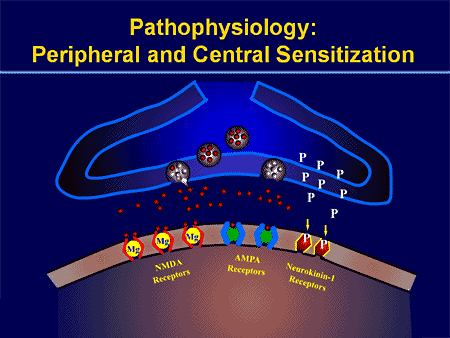
PERIFÉRNE MECHANIZMY
Po lézii periférneho nervu sa neuróny stávajú citlivejšími a rozvíja sa u nich abnormálna excitabilita a zvýšená citlivosť na stimuláciu
Toto je známe ako... Periférna senzibilizácia!
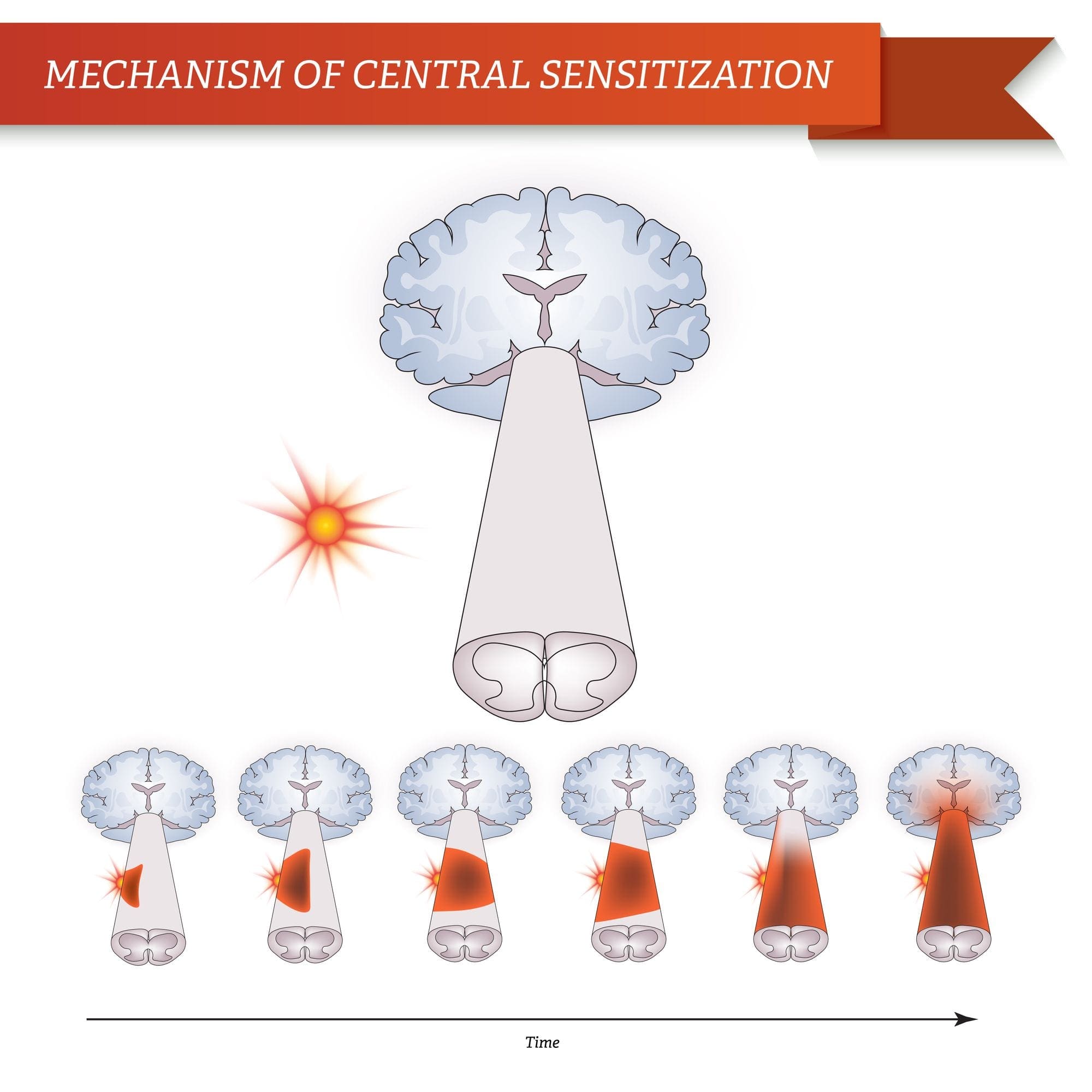
CENTRÁLNE MECHANIZMY
V dôsledku prebiehajúcej spontánnej aktivity vznikajúcej na periférii sa v neurónoch vyvíja zvýšená aktivita pozadia, zväčšené receptívne polia a zvýšené reakcie na aferentné impulzy, vrátane normálnych hmatových podnetov.
Toto je známe ako…Centrálna senzibilizácia!
BEŽNÉ PRÍČINY
Lézie alebo ochorenia somatosenzorického nervového systému môžu viesť k zmenenému a neusporiadanému prenosu senzorických signálov do miechy a mozgu; bežné stavy spojené s neuropatickou bolesťou zahŕňajú:
Postherpetická neuralgia
Neuralgia trojklanného nervu
Bolestivá radikulopatia
Diabetická neuropatia
HIV infekcia
malomocenstvo
amputácia
Bolesť pri poranení periférnych nervov
Mŕtvica (vo forme centrálnej bolesti po mŕtvici)
FANTÓMOVÁ BOLESŤ KONČATINY A ROZŠÍRENÁ REALITA
Fantomová bolesť končatín a AR
NEUROGÉNNY ZÁPAL
ciele:
Čo je to?
Aká je za tým patofyziológia?
Aké sú príčiny
Ako to môžeme opraviť?
NEUROGÉNNY ZÁPAL
Neurogénny zápal je nervovo vyvolaná lokálna zápalová odpoveď charakterizovaná vazodilatáciou, zvýšenou vaskulárnou permeabilitou, degranuláciou žírnych buniek a uvoľňovaním neuropeptidov vrátane SP a peptidu súvisiaceho s génom kalcitonínu (CGRP)
Zdá sa, že hrá dôležitú úlohu v patogenéze mnohých chorôb vrátane migrény, psoriázy, astmy, fibromyalgie, ekzému, rosacey, dystónie a mnohonásobnej chemickej citlivosti.
BEŽNÉ PRÍČINY
Existuje viacero ciest, ktorými môže byť iniciovaný neurogénny zápal. Je dobre zdokumentované s použitím zvieracích modelov aj izolovaných neurónov in vitro, že kapsaicín, teplo, protóny, bradykinín a tryptáza sú upstream regulátory intracelulárneho prítoku vápnika, čo vedie k uvoľňovaniu zápalového neuropeptidu. Na rozdiel od toho sa predpokladá, že prostaglandíny E2 a I2, cytokíny, interleukín-1, interleukín-6 a tumor nekrotizujúci faktor nespôsobujú samotné uvoľnenie neurotransmiterov, ale skôr excitujú senzorické neuróny, a tak znižujú prah pre spustenie a spôsobujú zvýšené uvoľňovanie neuropeptidy.
Zatiaľ čo neurogénny zápal bol rozsiahlo študovaný a dobre zdokumentovaný v periférnych tkanivách, donedávna zostal koncept neurogénneho zápalu v CNS do značnej miery nepreskúmaný. Vzhľadom na schopnosť neurogénneho zápalu ovplyvňovať vaskulárnu permeabilitu a viesť ku vzniku edému sa teraz široko skúmal jeho potenciál ovplyvniť permeabilitu BBB a vazogénny edém v mozgu a mieche za rôznych patologických podmienok.
Všetci nás ako deti učili, že existuje 5 zmyslov: zrak, chuť, zvuk, čuch a hmat. Počiatočné štyri zmysly využívajú jasné, zreteľné orgány, ako sú oči, chuťové poháriky, uši a nos, ale ako presne sa telesné zmysly dotýkajú? Dotyk sa prejavuje celým telom, zvnútra aj zvonka. Neexistuje jediný orgán, ktorý by bol zodpovedný za vnímanie dotyku. V celom tele sú skôr drobné receptory alebo nervové zakončenia, ktoré snímajú dotyk tam, kde k nemu dochádza, a vysielajú signály do mozgu s informáciami o type dotyku, ku ktorému došlo. Ako chuťový pohárik na jazyku deteguje chuť, mechanoreceptory sú žľazy v koži a na iných orgánoch, ktoré detegujú pocity dotyku. Sú známi ako mechanoreceptory pretože sú navrhnuté tak, aby zisťovali mechanické vnemy alebo rozdiely v tlaku.
Úloha mechanoreceptorov
Človek pochopí, že zažil pocit, keď orgán zodpovedný za objav tohto špecifického zmyslu pošle správu do mozgu, ktorý je primárnym orgánom, ktorý spracováva a usporiada všetky informácie. Správy sa posielajú zo všetkých oblastí tela do mozgu cez drôty označované ako neuróny. Existujú tisíce malých neurónov, ktoré sa rozvetvujú do všetkých oblastí ľudského tela a na koncoch mnohých z týchto neurónov sú mechanoreceptory. Aby sme demonštrovali, čo sa stane, keď sa dotknete predmetu, použijeme príklad.
Predstavte si, že vám na ruke pristane komár. Kmeň tohto hmyzu, taký ľahký, stimuluje mechanoreceptory v tejto konkrétnej oblasti ramena. Tieto mechanoreceptory posielajú správu pozdĺž neurónu, ku ktorému sú pripojené. Neurón sa pripojí až k mozgu, ktorý dostane správu, že sa niečo dotýka vášho tela v presnej polohe špecifického mechanoreceptora, ktorý správu vyslal. Mozog bude konať podľa tejto rady. Možno to povie očiam, aby sa pozreli na oblasť ruky, ktorá detekovala podpis. A keď oči povedia mozgu, že na paži je komár, mozog môže povedať ruke, aby ho rýchlo švihla preč. Tak fungujú mechanoreceptory. Účelom nižšie uvedeného článku je demonštrovať, ako aj podrobne diskutovať o funkčnej organizácii a molekulárnych determinantoch mechanoreceptorov.
Touch Sense: Funkčná organizácia a molekulárne determinanty mechanosenzitívnych receptorov
abstraktné
Kožné mechanoreceptory sú lokalizované v rôznych vrstvách kože, kde detegujú široké spektrum mechanických podnetov, vrátane ľahkého štetca, natiahnutia, vibrácií a škodlivého tlaku. Tejto rozmanitosti stimulov zodpovedá rôznorodá škála špecializovaných mechanoreceptorov, ktoré reagujú na kožnú deformáciu špecifickým spôsobom a prenášajú tieto stimuly do vyšších mozgových štruktúr. Štúdie mechanoreceptorov a geneticky ovládateľných zmyslových nervových zakončení začínajú odhaľovať mechanizmy vnímania dotyku. Práca v tejto oblasti poskytla výskumníkom dôkladnejšie pochopenie organizácie obvodov, ktoré sú základom vnímania dotyku. Nové iónové kanály sa objavili ako kandidáti na transdukčné molekuly a vlastnosti mechanicky hradlových prúdov zlepšili naše chápanie mechanizmov adaptácie na hmatové podnety. Tento prehľad zdôrazňuje pokrok dosiahnutý pri charakterizácii funkčných vlastností mechanoreceptorov v chlpatej a lysej koži a iónových kanáloch, ktoré detegujú mechanické vstupy a tvarujú adaptáciu mechanoreceptorov.
Kľúčové slová:mechanoreceptor, mechanosenzitívny kanál, bolesť, koža, somatosenzorický systém, dotyk
úvod
Dotyk je detekcia mechanického stimulu dopadajúceho na kožu, vrátane neškodných a škodlivých mechanických stimulov. Je to nevyhnutný zmysel pre prežitie a vývoj cicavcov a ľudí. Kontakt pevných predmetov a tekutín s pokožkou poskytuje potrebné informácie centrálnemu nervovému systému, ktorý umožňuje skúmanie a rozpoznávanie prostredia a iniciuje lokomóciu alebo plánovaný pohyb ruky. Dotyk je veľmi dôležitý aj pre učňovské vzdelanie, sociálne kontakty a sexualitu. Hmat je najmenej zraniteľným zmyslom, hoci pri mnohých patologických stavoch môže byť skreslený (hyperestézia, hypoestézia).1-3
Dotykové reakcie zahŕňajú veľmi presné kódovanie mechanických informácií. Kožné mechanoreceptory sú lokalizované v rôznych vrstvách kože, kde detegujú široké spektrum mechanických podnetov, vrátane ľahkého štetca, naťahovania, vibrácií, vychyľovania vlasov a škodlivého tlaku. Tejto rozmanitosti stimulov zodpovedá rôznorodá škála špecializovaných mechanoreceptorov, ktoré reagujú na kožnú deformáciu špecifickým spôsobom a prenášajú tieto stimuly do vyšších mozgových štruktúr. Somatosenzorické neuróny kože spadajú do dvoch skupín: nízkoprahové mechanoreceptory (LTMR), ktoré reagujú na benígny tlak, a vysokoprahové mechanoreceptory (HTMR), ktoré reagujú na škodlivú mechanickú stimuláciu. Bunkové telá LTMR a HTMR sa nachádzajú v dorzálnych koreňových gangliách (DRG) a kraniálnych senzorických gangliách (gangliá trojklaného nervu). Nervové vlákna spojené s LTMR a HTMR sú klasifikované ako Ap-, Ap- alebo C- vlákna na základe ich rýchlosti vedenia akčného potenciálu. C vlákna sú nemyelinizované a majú najnižšiu rýchlosť vedenia (~2 m/s), zatiaľ čo A? a A? vlákna sú ľahko a silne myelinizované, vykazujú strednú (~12 m/s) a rýchlu (~20 m/s) rýchlosť vedenia. LTMR sú tiež klasifikované ako pomaly alebo rýchlo sa adaptujúce reakcie (SA- a RA-LTMR) podľa ich rýchlosti adaptácie na trvalý mechanický stimul. Ďalej sa rozlišujú podľa kožných koncových orgánov, ktoré inervujú, a ich preferovaných stimulov.
Schopnosť mechanoreceptorov detegovať mechanické podnety závisí od prítomnosti iónových kanálov mechanoprevodníkov, ktoré rýchlo transformujú mechanické sily na elektrické signály a depolarizujú receptívne pole. Táto lokálna depolarizácia, nazývaná receptorový potenciál, môže vytvárať akčné potenciály, ktoré sa šíria smerom k centrálnemu nervovému systému. Vlastnosti molekúl, ktoré sprostredkúvajú mechanotransdukciu a adaptáciu na mechanické sily, však zostávajú nejasné.
V tomto prehľade poskytujeme prehľad vlastností mechanoreceptorov u cicavcov pri neškodnom a škodlivom dotyku chlpatej a lysej kože. Berieme do úvahy aj nedávne poznatky o vlastnostiach mechanicky hradlových prúdov v snahe vysvetliť mechanizmus adaptácie mechanoreceptorov. Nakoniec preskúmame nedávny pokrok dosiahnutý pri identifikácii iónových kanálov a súvisiacich proteínov zodpovedných za generovanie mechano-govaných prúdov.
Neškodný dotyk
LTMR spojené s vlasovým folikulom
Vlasové folikuly predstavujú miniorgány produkujúce vlasový hriadeľ, ktoré detegujú ľahký dotyk. Vlákna spojené s vlasovými folikulmi reagujú na pohyb vlasov a ich smer spúšťaním sledov akčných potenciálov pri nástupe a odstránení stimulu. Rýchlo sa prispôsobujú receptorom.
Mačka a králik. V srsti mačiek a králikov možno vlasové folikuly rozdeliť na tri typy vlasových folikulov, páperové vlasy, strážne srsti a tylotrichské. Dolné chlpy (spodná srsť, vlna, vellus)4 sú najpočetnejšie, najkratšie a najjemnejšie chlpy srsti. Sú zvlnené, bezfarebné a vychádzajú v skupinách po dvoch až štyroch chĺpkoch zo spoločného otvoru v koži. Vlásky strážcov (monotrichy, overhears, tophair)4 sú mierne zakrivené, buď pigmentované alebo nepigmentované, a vychádzajú jednotlivo z úst ich folikulov. Najmenej početné sú tielotrichy, najdlhšie a najhrubšie chĺpky.5,6 Sú pigmentované alebo nepigmentované, niekedy oboje a vychádzajú jednotlivo z folikulu, ktorý je obklopený slučkou kapilárnych krvných ciev. Senzorické vlákna zásobujúce vlasový folikul sa nachádzajú pod mazovou žľazou a pripisujú sa A? alebo vláknami A?-LTMR.7
V tesnej blízkosti spodného vlasu, tesne pod úrovňou mazovej žľazy, je prstenec kopijovitých pilo-Ruffini zakončení. Tieto senzorické nervové zakončenia sú umiestnené v špirálovitom smere okolo vlasového drieku v spojivovom tkanive tvoriacom vlasový folikul. Vo vlasovom folikule sú tiež voľné nervové zakončenia, niektoré z nich tvoria mechanoreceptory. Často obklopujú krčnú oblasť tylotrichového folikulu dotykové krvinky (pozri lysú kožu).
Vlastnosti myelinizovaných nervových zakončení v chlpatej koži mačiek a králikov boli intenzívne skúmané v období rokov 1930 – 1970 (prehľad v Hamann, 1995).8 Pozoruhodné je, že Brown a Iggo študovali 772 jednotiek s myelinizovanými aferentnými nervovými vláknami v safénových nervoch z mačiek. a králik, majú klasifikované reakcie v troch typoch receptorov, ktoré zodpovedajú pohybom chlpov Downa (receptory typu D), chlpov Guard (receptory typu G) a chlpov Tylotrichova (receptor typu T).9 Všetky reakcie aferentných nervových vlákien boli spojené v rýchlo sa adaptujúcom receptore typu I (RA I) opozíciou k Paciniánskemu receptoru s názvom RA II. Mechanoreceptory RA I detekujú rýchlosť mechanického stimulu a majú ostré hranice. Nezisťujú teplotné výkyvy. Burgess a kol. tiež opísali rýchlo sa adaptujúci receptor poľa, ktorý optimálne reaguje na hladenie pokožky alebo pohyb niekoľkých chĺpkov, čo sa pripisuje stimulácii pilo-Ruffiniho zakončení. Žiadna z odozvy vlasového folikulu sa nepripisovala aktivite vlákna C.10
Myšou. Na chrbtovej chlpatej koži myší boli opísané tri hlavné typy vlasových folikulov: cik-cak (okolo 72 %), šidlo/auchene (okolo 23 %) a chránič alebo tylotrich (okolo 5 %).11-14 Kľukaté a šidlo/ Auchenne vlasové folikuly produkujú tenšie a kratšie vlasové stonky a sú spojené s jednou mazovou žľazou. Guard alebo tylotrich vlasy sú najdlhšie z typov vlasových folikulov. Vyznačujú sa veľkou vlasovou cibuľkou spojenou s dvoma mazovými žľazami. Ochranné chĺpky a chĺpky šidla/auchene sú usporiadané v opakovanom, pravidelne rozmiestnenom vzore, zatiaľ čo cik-cak chĺpky husto obývajú oblasti pokožky obklopujúce dva väčšie typy vlasových folikulov [obr. 1 (A1, A2 a A3)].
Obrázok 1. Organizácia a projekcie kožných mechanoreceptorov. Pri ochlpenej pokožke sa ľahká kefa a dotyk detegujú najmä inerváciou okolo vlasových folikulov: šidlo/auchenne (A1), cikcak (A2) a chránič (A3). Chĺpky šidla/auchene sú trojnásobne inervované kopijovitými zakončeniami C-LTMR (A4), Ap-LTMR a A? rýchlo sa adaptujúci-LTMR (A6). Cik-cak vlasové folikuly sú kratšie vlasové stonky a sú inervované C-LTMR (A4) aj A? -LTMR kopijovité koncovky (A5). Najdlhšie ochranné vlasové folikuly sú inervované A? rýchlo sa adaptujúce-LTMR pozdĺžne kopijovité zakončenia (A6) a sú spojené s A? pomaly sa prispôsobujúce-LTMR koncoviek s dotykovou kupolou (A7). Centrálne výbežky všetkých týchto vlákien končia v odlišných, ale čiastočne sa prekrývajúcich laminách chrbtového rohu miechy (C-LTMR v lamine II, Ap-LTMR v lamine III a Ap-LTMR v lamine IV a V). Projekcie LTMR, ktoré inervujú rovnaké alebo susedné vlasové folikuly, sú zarovnané tak, aby vytvorili úzky stĺpec v dorzálnom rohu miechy (B1 v sivej farbe). Len pri ochlpenej pokožke subpopulácia voľných zakončení C-vlákien inervuje epidermis a reaguje na príjemný dotyk (A8). Tieto vlákna C-touch nereagujú na škodlivý dotyk a ich dráha pohybu zatiaľ nie je známa (B2). V lysej koži je neškodný dotyk sprostredkovaný štyrmi typmi LTMR. Komplex Merkelových buniek a neuritov sa nachádza v bazálnej vrstve epidermis (C1). Tento mechanoreceptor pozostáva z usporiadania medzi mnohými Merkelovými bunkami a zväčšeným nervovým zakončením z jedného A? vlákno. Merkelove bunky vykazujú procesy podobné prstom, ktoré sú v kontakte s keratinocytmi (C2). Ruffiniho zakončenie je lokalizované v derme. Ide o tenké zapuzdrené senzorické zakončenia v tvare cigary spojené s A? vlákno (C3). Meissnerove telieska spojené s A? nervové zakončenie a nachádza sa v dermálnych papilách. Tento zapuzdrený mechanoreceptor pozostáva zo zbalených podporných buniek usporiadaných ako horizontálne lamely obklopené spojivovým tkanivom (C4). Paciniánske teliesko je hlbší mechanoreceptor. Jedno jediné A? nemyelinizované nervové zakončenie končí v strede tohto veľkého vajcovitého telieska tvoreného koncentrickými lamelami. Projekcie týchto vlákien Ap-LTMR v mieche sú rozdelené do dvoch vetiev. Hlavná centrálna vetva (B3) stúpa v mieche v ipsilaterálnych dorzálnych formujúcich klinovitých alebo gracilových zväzkoch (B5) na úrovni drene, kde primárne aferenty vytvárajú svoju prvú synapsiu (B6). Sekundárne neuróny vykonajú senzorickú dekusáciu (B7), aby vytvorili trakt na mediálnom lemnisku, ktorý stúpa cez mozgový kmeň do stredného mozgu, konkrétne do talamu. Sekundárna vetva LTMR končí v dorzálnom rohu v lamine II, IV, V a interferuje s prenosom bolesti (B4). Škodlivý dotyk je detekovaný voľným nervovým zakončením v epidermis chlpatej (A9) aj lysej kože (C7). Tieto mechanoreceptory sú zakončením Ap-HTMR a C-HTMR v tesnom kontakte so susednými keratinocytmi (C6). Ap-hTMR končí v lamine I a V; C-HTMR končí v lamine I a II (B8). Na úrovni chrbtového rohu miechy vytvárajú primárne aferentné HTMR synapsie so sekundárnymi neurónmi, ktoré prechádzajú strednou čiarou a stúpajú do vyššej mozgovej štruktúry v anterolaterálnom zväzku (B9, B10).
Ginty a spolupracovníci nedávno použili kombináciu molekulárno-genetického označovania a somatotopického retrográdneho sledovania na vizualizáciu organizácie periférnych a centrálnych axonálnych zakončení LTMR u myší.15 Ich zistenia podporujú model, v ktorom sú jednotlivé znaky komplexného hmatového stimulu extrahované tromi typmi vlasových folikulov a prenášané prostredníctvom aktivít jedinečných kombinácií Ap-, Ap- a C- vlákien do dorzálneho rohu.
Ukázali, že genetické značenie tyrozínhydroxylázy pozitívnych (TH+) DRG neurónov charakterizuje populáciu nepeptidergických senzorických neurónov s malým priemerom a umožňuje vizualizáciu periférnych zakončení C-LTMR v koži. Prekvapivo sa zistilo, že axoneálne vetvy jednotlivých C-LTMR sa arborizujú a tvoria pozdĺžne kopijovité zakončenia, ktoré sú úzko spojené s cikcakom (80 % zakončení) a šidlom/auchene (20 % zakončení), ale nie s tylotrichovými vlasovými folikulmi [obr. 1 (A4)]. Dlho sa predpokladalo, že pozdĺžne kopijovité zakončenia patria výlučne k A?-LTMR, a preto sa neočakávalo, že zakončenia C-LTMR budú tvoriť pozdĺžne kopijovité zakončenia.15 Tieto C-LTMR majú prechodnú adaptáciu v porovnaní s pomaly a rýchlo sa adaptujúcimi myelinizované mechanoreceptory [obr. 2 (Cl)].
Obrázok 2. Hmatové receptory u cicavcov: Kožné hmatové receptory sa diferencujú na neškodný dotyk podporovaný viacerými receptormi s nízkym mechanickým prahom (LTMR) v lysej a chlpatej koži a škodlivý dotyk podporovaný receptorom vysokého mechanického prahu (HTMR). Tvoria voľné nervové zakončenia, ktoré končia hlavne v epiderme. (A) Lysá koža. A1: Meissnerove telieska detegujú pohyb kože a skĺznutie predmetu v ruke. Sú dôležité pre manipuláciu s predmetmi a zručnosť. Receptory sa rýchlo prispôsobujú stimulu, sú spojené s A? vlákna a riedko na vlákna C a majú veľké receptorové pole. Odpoveď 2: Korpuskuly Ruffini detekujú natiahnutie kože a sú dôležité na zistenie polohy prstov a podávaného predmetu. Receptor sa pomaly prispôsobuje stimulu a udržiava aktivitu tak dlho, ako je stimul aplikovaný. Receptory sú pripojené k A? vlákna a majú veľké receptívne pole. A3: Paciniánske telieska sú hlbšie v derme a detegujú vibrácie. Receptory sú pripojené k A? vlákna; rýchlo sa prispôsobujú podnetu a majú najväčšie receptívne pole. (B) Celá koža. B1: Komplexy Merkelových buniek sú prítomné v lysej koži aj okolo vlasov. Sú husto vyjadrené v ruke a sú dôležité pre vnímanie textúry a najjemnejšie rozlíšenie medzi dvoma bodmi. Sú zodpovedné za presnosť prstov. Receptory sú pripojené k A? vlákna; pomaly sa prispôsobujú podnetu a majú krátke receptívne pole. B2: Škodlivé dotykové HTMR s veľmi pomalou adaptáciou na stimul, tj aktívne, pokiaľ je aplikovaný nociceptívny stimul. Sú tvorené voľným nervovým zakončením A? a C-vlákna spojené s keratinocytmi. (C) Chlpatá pokožka. C1: Vlasové folikuly sú spojené s rôznymi typmi vlasov. U myší sú chlpy Guard dlhšie a riedko vyjadrené, šidlo/auchenne sú stredne veľké a cik-cak sú najmenšie a najhustejšie vyjadrené chlpy. Sú pripojené k A? vlákna ale aj do A? a C-LTMR vlákna pre šidlo/auchenne a cikcakové vlasy. Detekujú pohyb vlasov vrátane príjemného dotyku počas maznania. Rýchlo alebo so strednou kinetikou sa adaptujú na stimul. C2: C-dotykové nervové zakončenia zodpovedajú podtypu zakončenia C vlákien s voľným zakončením charakterizovaným nízkym mechanickým prahom. Majú zakódovať príjemný pocit vyvolaný pohladením. Mierne sa prispôsobujú podnetom a majú krátke receptívne pole. Predpokladané mechanosenzitívne (MS) iónové kanály vyjadrené v rôznych hmatových receptoroch sú označené podľa predbežných údajov a sumarizujú súčasnú vyhodnocovanú hypotézu.
Druhá hlavná identifikovaná populácia sa týka Ap-LTMR zakončení v Awl/Auchenne a cikcak folikuloch, ktoré majú byť porovnané s Downovým vlasovým folikulom, ktorý sa intenzívne študoval u mačky a králika. Ginty a spolupracovníci ukázali, že TrkB je exprimovaný vo vysokých hladinách v podskupine neurónov DRG so stredným priemerom. Intracelulárne záznamy s použitím ex vivo preparátu kožných nervov značených vlákien odhalili, že vykazujú fyziologické vlastnosti vlákien predtým študovaných u mačiek a králikov: vynikajúca mechanická citlivosť (Von Freyov prah < 0.07 mN), rýchle prispôsobenie sa odozvy na nadprahové podnety, stredné vedenie rýchlosti (5.8 � 0.9 m/s) a úzke neohnuté soma hroty.15 Tieto A?-LTMR tvoria pozdĺžne kopijovité zakončenia spojené prakticky s každým cikcakovitým a šidlom/auchene vlasovým folikulom trupu [obr. 1 (A5)].
Nakoniec ukázali, že periférne zakončenia rýchlo sa adaptujúceho A? LTMR tvoria pozdĺžne kopijovité zakončenia spojené s ochrannými (alebo tylotrichovými) a šidlovými/auchenovými vlasovými folikulmi [obr. 1 (A6)].15 Okrem toho sú chĺpky strážcov spojené aj s komplexom Merkelových buniek tvoriacim dotykovú kupolu spojenú s A? pomaly sa adaptujúci LTMR [obr. 1 (A7)].
Stručne povedané, prakticky všetky cik-cak vlasové folikuly sú inervované kopijovitými zakončeniami C-LTMR aj Ap-LTMR; chĺpky šidla/auchene sú trojnásobne inervované A? rýchlo sa adaptujúce-LTMR, Ap-LTMR a C-LTMR kopijovité zakončenia; Ochranné vlasové folikuly sú inervované A? rýchlo sa adaptujúce-LTMR pozdĺžne kopijovité zakončenia a interagujú s A? pomaly sa prispôsobujúci-LTMR dotykových kupolových zakončení. Každý myší vlasový folikul teda dostáva jedinečné a nemenné kombinácie zakončení LTMR zodpovedajúcich neurofyziologicky odlišným mechanosenzorickým koncovým orgánom. Vzhľadom na iteratívne usporiadanie týchto troch typov vlasov Ginty a spolupracovníci navrhujú, aby chlpatá pokožka pozostávala z iteratívneho opakovania periférnej jednotky obsahujúcej (1) jeden alebo dva centrálne umiestnené ochranné vlasy, (2) ~20 okolitých chĺpkov šidla/auchenne a (3 ) ~80 roztrúsených cik-cak vlasov [obr. 2 (Cl)].
Projekcia miechy. Centrálne projekcie A? rýchlo sa adaptujúce LTMR, Ap-LTMR a C-LTMR končia v odlišných, ale čiastočne sa prekrývajúcich laminách (II, III, IV) dorzálneho rohu miechy. Okrem toho sú centrálne terminály LTMR, ktoré inervujú rovnaké alebo susedné vlasové folikuly v rámci periférnej jednotky LTMR, zarovnané tak, aby vytvorili úzky stĺpec LTMR v dorzálnom rohu miechy [obr. 1 (B1)]. Zdá sa teda pravdepodobné, že klin alebo stĺpec somatotopicky organizovaných primárnych senzorických aferentných zakončení v dorzálnom rohu predstavuje zarovnanie centrálnych projekcií Ap-, Ap- a C-LTMR, ktoré inervujú rovnakú periférnu jednotku a detegujú mechanické stimuly pôsobiace na rovnakú malú skupinu vlasových folikulov. Ginty a spolupracovníci odhadujú, že na základe počtu chlpov na trupe a končatinách chlpov na šidle, šidlách a cikcakovitých chlpoch na trupe a končatinách a na počtoch každého podtypu LTMR, Ginty a spolupracovníci odhadujú, že chrbtový roh myši obsahuje 2,000 4,000 – 15 XNUMX stĺpcov LTMR, čo zodpovedá približnému počtu periférnych Jednotky LTMR.XNUMX
Okrem toho sú axóny podtypov LTMR navzájom úzko spojené, majú prepletené výbežky a vzájomne prepojené kopijovité zakončenia, ktoré inervujú rovnaký vlasový folikul. Okrem toho, pretože tieto tri typy vlasových folikulov vykazujú rôzne tvary, veľkosti a bunkové zloženie, je pravdepodobné, že budú mať odlišné ohybové alebo vibračné ladiace vlastnosti. Tieto zistenia sú v súlade s klasickými neurofyziologickými meraniami u mačky a králika, čo naznačuje, že A? RA-LTMR a Ap-LTMR môžu byť rozdielne aktivované vychýlením rôznych typov vlasových folikulov.16,17
Na záver, dotyk v chlpatej koži je kombináciou: (1) relatívnych počtov, jedinečných priestorových distribúcií a odlišných morfologických a ohybových vlastností troch typov vlasových folikulov; (2) jedinečné kombinácie koncoviek podtypu LTMR spojené s každým z troch typov vlasových folikulov; a (3) rozdielne citlivosti, rýchlosti vedenia, vzory hrotov a adaptačné vlastnosti štyroch hlavných tried LTMR spojených s vlasovými folikulami, ktoré umožňujú mechanosenzorickému systému chlpatej kože extrahovať a preniesť do CNS komplexné kombinácie vlastností, ktoré definujú dotyk.
LTMR s voľnými nervami
Vo všeobecnosti sú konce C-vlákien v koži HTMR, ale subpopulácia C-vlákien nereaguje na škodlivý dotyk. Táto podskupina hmatových aferentných vlákien C (CT) predstavuje odlišný typ nemyelinizovaných, nízkoprahových mechanoreceptívnych jednotiek existujúcich vo chlpatej, ale nie lysej koži ľudí a cicavcov [obr. 1 (A8)].18,19 CT sa vo všeobecnosti spájajú s vnímaním príjemnej hmatovej stimulácie pri telesnom kontakte.20,21
CT aferenty reagujú na vtlačovacie sily v rozsahu 0.3 – 2.5 mN a sú teda rovnako citlivé na deformáciu kože ako mnohé z A? Adaptačné charakteristiky CT aferentov sú teda stredné v porovnaní s pomaly a rýchlo sa adaptujúcimi myelinizovanými mechanoreceptormi. Recepčné polia ľudských CT aferentácií majú zhruba okrúhly alebo oválny tvar. Pole pozostáva z jedného až deviatich malých citlivých škvŕn rozmiestnených na ploche do 19 mm35 Myšie homológne receptory sú organizované vo vzore nesúvislých škvŕn pokrývajúcich asi 2.22 – 50 % plochy chlpatej kože [obr. 60 (C2)].2
Dôkazy od pacientov bez myelinizovaných hmatových aferentov naznačujú, že signalizácia v CT vláknach aktivuje ostrovnú kôru. Keďže tento systém je slabý v kódovaní diskriminačných aspektov dotyku, ale dobre sa hodí na kódovanie pomalého, jemného dotyku, CT vlákna vo chlpatej koži môžu byť súčasťou systému na spracovanie príjemných a spoločensky relevantných aspektov dotyku.24 Aktivácia CT vlákna môže tiež majú úlohu pri inhibícii bolesti a nedávno sa navrhlo, že zápal alebo trauma môžu zmeniť vnem prenášaný LTMR s C-vláknom z príjemného dotyku na bolesť.25,26
Ktorá dráha CT-aferentných ciest zatiaľ nie je známa [obr. 1 (B2)], ale boli zdokumentované nízkoprahové hmatové vstupy do spinothalamických projekčných buniek27, čo potvrdzuje dôveryhodnosť správ o jemných kontralaterálnych deficitoch detekcie dotyku u ľudských pacientov po deštrukcii týchto dráh po chordotómii.28
LTMR v lysej koži
Komplexy Merkelových buniek a neuritov a dotyková kupola. Merkelová (1875) ako prvá podala histologický popis zhlukov epidermálnych buniek s veľkými laločnatými jadrami, ktoré nadväzujú kontakt s predpokladanými aferentnými nervovými vláknami. Predpokladal, že si podriaďujú hmat tým, že ich nazývajú Tastzellen (hmatové bunky). U ľudí sú komplexy Merkelových buniek obohatené v oblastiach citlivých na dotyk, nachádzajú sa v bazálnej vrstve epidermis na prstoch, perách a genitáliách. Vyskytujú sa aj v chlpatej koži v nižšej hustote. Komplex Merkelovej bunky a neuritov pozostáva z Merkelovej bunky v tesnej blízkosti zväčšeného nervového zakončenia z jedného myelinizovaného A? vlákno [obr. 1 (C1)] (prehľad v Halata a spolupracovníci).29 Na epidermálnej strane vykazuje Merkelová bunka prstovité výbežky siahajúce medzi susednými keratinocytmi [obr. 1 (C2)]. Merkelove bunky sú epidermálne bunky odvodené od keratinocytov.30,31 Termín dotyková kupola bol zavedený na pomenovanie veľkej koncentrácie komplexov Merkelových buniek vo chlpatej koži prednej labky mačky. Dotyková kupola môže mať až 150 Merkelových buniek inervovaných jedným Ap-vláknom a u ľudí okrem Ap-vlákien môže A? a pravidelne boli prítomné aj C-vlákna.32-34
Stimulácia komplexov Merkelových buniek má za následok pomaly sa adaptujúce reakcie typu I (SA I), ktoré pochádzajú z prerušovaných receptívnych polí s ostrými hranicami. Nedochádza k spontánnemu výtoku. Tieto komplexy reagujú na hĺbku vtlačenia kože a majú najvyššie priestorové rozlíšenie (0.5 mm) z kožných mechanoreceptorov. Prenášajú presný priestorový obraz hmatových podnetov a navrhuje sa, aby boli zodpovedné za rozlišovanie tvaru a textúry [obr. 2 (B1)]. Myši bez Merkelových buniek nedokážu rozpoznať textúrované povrchy nohami, zatiaľ čo tak robia pomocou fúzov.35
Či sú Merkelová bunka, senzorický neurón alebo oboje miestami mechanotransdukcie, je stále predmetom diskusie. U potkanov fototoxická deštrukcia Merkelových buniek ruší SA I odpoveď.36 U myší s geneticky potlačenými Merkelovými bunkami SA I odpoveď zaznamenaná ex vivo prípravou kože/nervu úplne vymizla, čo dokazuje, že Merkelove bunky sú potrebné na správne kódovanie Merkelovej Avšak mechanická stimulácia izolovaných Merkelových buniek v kultúre motoricky riadeným tlakom negeneruje mechanicky riadené prúdy.37 Keratinocyty môžu hrať dôležitú úlohu v normálnom fungovaní komplexu Merkelových buniek a neuritov. Procesy podobné prstom Merkelovej bunky sa môžu pohybovať s deformáciou kože a pohybom buniek epidermis, a to môže byť prvý krok mechanickej transdukcie. Je zrejmé, že podmienky potrebné na štúdium mechanickej citlivosti Merkelových buniek ešte musia byť stanovené.
Ruffiniho zakončenia. Ruffini koncovky sú tenké zapuzdrené senzorické koncovky v tvare cigary spojené s A? nervových zakončení. Ruffini zakončenia sú malé valčeky spojivového tkaniva usporiadané pozdĺž dermálnych kolagénových vlákien, ktoré sú zásobované jedným až tromi myelinizovanými nervovými vláknami s priemerom 4�6�m. Až tri valce rôznej orientácie v derme sa môžu spojiť a vytvoriť jeden receptor [obr. 1 (C3)]. Štrukturálne sú zakončenia Ruffini podobné orgánom Golgiho šľachy. Sú široko exprimované v derme a boli identifikované ako pomaly sa adaptujúce kožné mechanoreceptory typu II (SA II). Na pozadí spontánnej nervovej aktivity je pomaly sa prispôsobujúci pravidelný výboj vyvolaný kolmou nízkou silou udržiavanou mechanickou stimuláciou alebo účinnejšie dermálnym naťahovaním. Odpoveď SA II pochádza z veľkých receptívnych polí s nejasnými hranicami. Ruffiniho receptory prispievajú k vnímaniu smeru pohybu objektu prostredníctvom vzoru natiahnutia kože [obr. 2 (A2)].
U myší sa reakcie SA I a SA II dajú oddeliť elektrofyziologicky v prípravku nerv-koža ex vivo.40 Nandasena a spolupracovníci uviedli imunolokalizáciu akvaporínu 1 (AQP1) v periodontálnych Ruffiniho zakončeniach rezákov potkana, čo naznačuje, že AQP1 sa podieľa na udržiavanie dentálnej osmotickej rovnováhy potrebnej pre mechanotransdukciu.41 Periodontálne Ruffiniho zakončenia tiež vyjadrovali predpokladaný mechanosenzitívny iónový kanál ASIC3.42
Meissnerove krvinky. Meissnerove telieska sú lokalizované v dermálnych papilách lysej kože, najmä v dlaniach a chodidlách, ale aj v perách, jazyku, tvári, bradavkách a genitáliách. Anatomicky pozostávajú zo zapuzdreného nervového zakončenia, pričom puzdro je vyrobené zo sploštených podporných buniek usporiadaných ako horizontálne lamely zapustené do spojivového tkaniva. Existuje jedno nervové vlákno A? aferenty spojené na telieska [obr. 1 (C4)]. Akákoľvek fyzická deformácia telieska spustí salvu akčných potenciálov, ktoré rýchlo ustanú, tj rýchlo sa adaptujú na receptory. Keď sa stimul odstráni, teliesko znovu nadobudne svoj tvar a pri tom vytvára ďalšiu salvu akčných potenciálov. Vďaka svojej povrchovej polohe v derme tieto krvinky selektívne reagujú na pohyb kože, hmatovú detekciu pošmyknutia a vibrácie (20�40 Hz). Sú citlivé na dynamickú pokožku – napríklad medzi pokožkou a predmetom, s ktorým sa manipuluje [obr. 2 (A1)].
Pacinovské krvinky. Paciniánske telieska sú hlbšie mechanoreceptory kože a sú najcitlivejším zapuzdreným kožným mechanoreceptorom pohybu kože. Tieto veľké vajcovité telieska (dĺžka 1 mm) vyrobené zo sústredných lamiel vláknitého spojivového tkaniva a fibroblastov vystlaných plochými modifikovanými Schwannovými bunkami sú exprimované v hlbokej dermis.43 V strede telieska, v dutine naplnenej tekutinou, nazývanej vnútorný bulbus , končí jedno jediné A? aferentné nemyelinizované nervové zakončenie [obr. 1 (C5)]. Majú veľké receptívne pole na povrchu pokožky s obzvlášť citlivým stredom. U mutantných myší c-Maf je narušený vývoj a funkcia niekoľkých rýchlo sa adaptujúcich typov mechanoreceptorov. Najmä Pacinove telieska sú silne atrofované.44
Pacinove telieska vykazujú veľmi rýchlu adaptáciu ako odpoveď na vrúbkovanie kože, rýchlo sa adaptujúci nervový výboj II (RA II), ktorý je schopný sledovať vysokú frekvenciu vibračných stimulov a umožňuje vnímanie vzdialených udalostí prostredníctvom prenášaných vibrácií.45 Pacinove telieska aferenty reagujú na trvalú indentáciu s prechodnou aktivitou pri nástupe a odsadení stimulu. Nazývajú sa tiež detektory zrýchlenia, pretože dokážu zistiť zmeny v sile stimulu a ak sa rýchlosť zmeny stimulu zmení (ako sa to deje pri vibráciách), ich odozva sa stane úmernou tejto zmene. Pacinove telieska vnímajú hrubé zmeny tlaku a predovšetkým vibrácie (150�300 Hz), ktoré dokážu rozpoznať aj na centimetre [obr. 2 (A3)].
Tonická odpoveď bola pozorovaná v dekapsulovanom Pacinovom teliesku.46 Okrem toho intaktné Pacinove telieska reagujú trvalou aktivitou počas konštantných indentačných stimulov, bez zmeny mechanických prahov alebo frekvencie odozvy, keď je signalizácia sprostredkovaná GABA blokovaná medzi lamelovou gliou a nervovým zakončením.47 neneurónové zložky Pacinovho telieska môžu mať dvojitú úlohu pri filtrovaní mechanického stimulu, ako aj pri modulácii vlastností odozvy senzorického neurónu.
Projekcie miechy. Projekcie Ap-LTMR v mieche sú rozdelené do dvoch vetiev. Hlavná centrálna vetva stúpa v mieche v ipsilaterálnych chrbtových stĺpcoch na cervikálnu úroveň [obr. 1 (B3)]. Sekundárne vetvy končia v dorzálnom rohu v laminách IV a interferujú napríklad s prenosom bolesti. To môže zmierniť bolesť ako súčasť ovládania brány [obr. 1 (B4)].48
Na cervikálnych úrovniach sa axóny hlavnej vetvy oddeľujú na dva trakty: trakt strednej čiary obsahuje gracilný zväzok prenášajúci informácie z dolnej polovice tela (nohy a trup) a vonkajší trakt obsahuje klinovitý zväzok prenášajúci informácie z hornej polovice tela (ruky a trup) [obr. 1 (B5)].
Primárne hmatové aferenty vytvárajú svoju prvú synapsiu s neurónmi druhého rádu v dreni, kde vlákna z každého traktu synapsujú v jadre rovnakého mena: synapsia axónov gracile fasciculus v jadre gracile a synapsia klinovitých axónov v jadre klinovitého tvaru [obr. 1 (B6)]. Neuróny prijímajúce synapsiu poskytujú sekundárne aferenty a okamžite prechádzajú strednou čiarou, aby vytvorili trakt na kontralaterálnej strane mozgového kmeňa – mediálny lemniscus – ktorý stúpa cez mozgový kmeň do ďalšej prenosovej stanice v strednom mozgu, konkrétne v talame [obr. . 1 (B7)].
Molekulárna špecifikácia LTMR. Molekulárne mechanizmy kontrolujúce skorú diverzifikáciu LTMR boli nedávno čiastočne objasnené. Bourane a spolupracovníci ukázali, že neurónové populácie exprimujúce Ret tyrozínkinázový receptor (Ret) a jeho koreceptor GFR?2 u E11�13 embryonálnych myší DRG selektívne koexprimujú transkripčný faktor Mafa.49,50 Títo autori demonštrujú, že Mafa/ Ret/GFRy2 neuróny predurčené stať sa tromi špecifickými typmi LTRM pri narodení: SA1 neuróny inervujúce komplexy Merkelových buniek, rýchlo sa adaptujúce neuróny inervujúce Meissnerove telieska a rýchlo sa adaptujúce aferenty (RA I) tvoriace kopijovité zakončenia okolo vlasových folikulov. Ginty a spolupracovníci tiež uvádzajú, že DRG neuróny exprimujúce skorý Ret sa rýchlo prispôsobujú mechanoreceptorom z Meissnerových teliesok, Paciniových teliesok a kopijovitých zakončení okolo vlasových folikulov.51 Inervujú diskrétne cieľové zóny v gracile a klinovité jadrá, čo odhaľuje špecifický vzorec modality. neurónové axonálne projekcie v mozgovom kmeni.
Prieskum mechanoreceptorov ľudskej kože. Technika �mikroneurografie�, ktorú opísali Hagbarth a Vallbo v roku 1968, bola aplikovaná na štúdium výbojového správania jednotlivých ľudských mechanosenzitívnych zakončení zásobujúcich svaly, kĺby a kožu (pozri prehľad Macefield, 2005).52,53 Väčšina mikroneurografie ľudskej kože štúdie charakterizovali fyziológiu hmatových aferentných kanálov v lysej koži ruky. Mikroelektródové záznamy zo stredných a ulnárnych nervov u ľudských subjektov odhalili dotykový pocit generovaný štyrmi triedami LTMR: Meissnerove aferenty sú obzvlášť citlivé na ľahké ťahanie cez kožu, reagujúc na lokálne šmykové sily a začínajúce alebo zjavné sklzy v receptívnom poli. Paciniánske aferenty sú mimoriadne citlivé na rýchle mechanické prechody. Aferenty energicky reagujú na fúkanie cez receptívne pole. Pacinovské teliesko umiestnené v číslici zvyčajne reaguje na poklepanie na stôl podopierajúci rameno. Merkelove aferenty majú charakteristicky vysokú dynamickú citlivosť na indentačné stimuly aplikované na diskrétnu oblasť a často reagujú uvoľnením výboja počas uvoľňovania. Hoci Ruffiniho aferenty reagujú na sily pôsobiace normálne na kožu, jedinečnou vlastnosťou SA II aferentov je ich schopnosť reagovať aj na laterálne natiahnutie kože. Nakoniec, vlasové jednotky na predlaktí majú veľké vajcovité alebo nepravidelné receptívne polia zložené z viacerých citlivých miest, ktoré zodpovedali jednotlivým vlasom (každý aferentný dodáva ~ 20 vlasov).
Mechanická citlivosť keratinocytov
Akýkoľvek mechanický stimul na koži sa musí preniesť cez keratinocyty, ktoré tvoria epidermis. Tieto všadeprítomné bunky môžu okrem podporných alebo ochranných úloh vykonávať aj signalizačné funkcie. Napríklad keratinocyty vylučujú ATP, dôležitú senzorickú signálnu molekulu, v reakcii na mechanické a osmotické podnety.54,55 Uvoľňovanie ATP indukuje zvýšenie intracelulárneho vápnika autokrinnou stimuláciou purinergných receptorov.55 Okrem toho existujú dôkazy, že hypotonicita aktivuje Rho -kinázová signálna dráha a následná tvorba stresového vlákna F-aktínu, čo naznačuje, že mechanická deformácia keratinocytov môže mechanicky interferovať so susednými bunkami, ako sú Merkelove bunky pre neškodný dotyk a konce bez C-vlákna pre škodlivý dotyk [obr. 1 (C6)].56,57
Škodlivý dotyk
Vysokoprahové mechanoreceptory (HTMR) sú epidermálne C- a A? voľné nervové zakončenia. Nie sú spojené so špecializovanými štruktúrami a sú pozorované v oboch chlpatých kožiach [obr. 1 (A9)] a lysej kože [obr. 1(C7)]. Termín voľných nervových zakončení je však potrebné zvážiť obozretne, pretože nervové zakončenia sú vždy v tesnej blízkosti keratinocytov alebo Langheransových buniek alebo melanocytov. Ultraštrukturálna analýza nervových zakončení odhaľuje prítomnosť hrubého endoplazmatického retikula, hojné mitochondrie a vezikuly s hustým jadrom. Priľahlé membrány epidermálnych buniek sú zhrubnuté a pripomínajú postsynaptickú membránu v nervových tkanivách. Všimnite si, že interakcie medzi nervovými zakončeniami a epidermálnymi bunkami môžu byť obojsmerné, pretože epidermálne bunky môžu uvoľňovať mediátory ako ATP, interleukín (IL6, IL10) a bradykinín a naopak peptidergické nervové zakončenia môžu uvoľňovať peptidy, ako je CGRP alebo látka P pôsobiaca na epidermálne bunky. HTMR zahŕňajú mechano-nociceptory excitované iba škodlivými mechanickými stimulmi a polymodálne nociceptory, ktoré tiež reagujú na škodlivé teplo a exogénne chemikálie [obr. 2 (B2)].58
HTMR aferentné vlákna končia na projekčných neurónoch v dorzálnom rohu miechy. Ap-HTMR kontaktujú neuróny druhého rádu prevažne v lamine I a V, zatiaľ čo C-HTMR končia v lamine II [obr. 1 (B8)]. Nociceptívne neuróny druhého rádu vyčnievajú na kontrolnú stranu miechy a stúpajú v bielej hmote, čím vytvárajú anterolaterálny systém. Tieto neuróny končia hlavne v talame [obr. 1 (B9 a B10)].
Mechanoprúdy v somatosenzorických neurónoch
Mechanizmy pomalej alebo rýchlej adaptácie mechanoreceptorov ešte nie sú objasnené. Nie je jasné, do akej miery je adaptácia mechanoreceptorov zabezpečená bunkovým prostredím senzorického nervového zakončenia, vnútornými vlastnosťami mechanicky riadených kanálov a vlastnosťami axonálnych napäťovo riadených iónových kanálov v senzorických neurónoch (obr. 2). Nedávny pokrok v charakterizácii mechanicky hradlovaných prúdov však ukázal, že v DRG neurónoch existujú rôzne triedy mechanosenzitívnych kanálov a môže vysvetliť niektoré aspekty adaptácie mechanoreceptorov.
In vitro záznam u hlodavcov ukázal, že sóma neurónov DRG je vnútorne mechanicky senzitívna a exprimuje katiónové mechanicky riadené prúdy.59-64 Gadolínium a ruténiová červeň úplne blokujú mechanosenzitívne prúdy, zatiaľ čo vonkajší vápnik a horčík vo fyziologických koncentráciách, ako aj amilorid a benzamil spôsobujú čiastočný blok.60,62,63 FM1-43 pôsobí ako trvalý blokátor a injekcia FM1-43 do zadnej labky myší znižuje citlivosť na bolesť v Randall-Selittovom teste a zvyšuje hodnotený prah stiahnutia labky s von Freyovými vlasmi.65
V reakcii na trvalú mechanickú stimuláciu mechanosenzitívne prúdy klesajú uzavretím. Na základe časových konštánt poklesu prúdu boli rozlíšené štyri odlišné typy mechanosenzitívnych prúdov: rýchlo sa prispôsobujúce prúdy (~ 3�6 ms), prechodne sa prispôsobujúce prúdy (~15�30 ms), pomaly sa prispôsobujúce prúdy (~200�300 ms ) a ultra-pomaly sa adaptujúce prúdy (~1000 ms).64 Všetky tieto prúdy sú prítomné s rôznym výskytom v potkaních DRG neurónoch inervujúcich lysú kožu zadnej labky.64
Mechanickú citlivosť mechanosenzitívnych prúdov možno určiť aplikáciou série prírastkových mechanických stimulov, čo umožňuje relatívne podrobnú analýzu stimul-prúd.66 Vzťah medzi stimulom a prúdom je typicky sigmoidálny a maximálna amplitúda prúdu je určená počtom kanály, ktoré sú súčasne otvorené.64,67 Zaujímavé je, že rýchlo sa adaptujúci mechanickosenzitívny prúd vykazuje nízky mechanický prah a strednú hodnotu polovičnej aktivácie v porovnaní s ultra pomaly sa adaptujúcim mechanosenzitívnym prúdom.63,65
Senzorické neuróny s nenociceptívnymi fenotypmi prednostne exprimujú rýchlo sa adaptujúce mechanosenzitívne prúdy s nižším mechanickým prahom.60,61,63,64,68 Naopak, pomaly a ultrapomaly sa adaptujúce mechanosenzitívne prúdy sú občas hlásené v domnelých nenociceptívnych bunkách.64,68 To vyvolalo návrh, že tieto prúdy môžu prispieť k rôznym mechanickým prahom pozorovaným v LTMR a HTMR in vivo. Hoci tieto in vitro experimenty by sa mali brať s opatrnosťou, podporu prítomnosti DRG neurónov v sóme nízkoprahových a vysokoprahových mechanotransduktorov poskytla aj stimulácia kultivovaných myších senzorických neurónov na báze radiálneho roztiahnutia.69 Táto paradigma odhalila dve hlavné populácie neurónov citlivých na natiahnutie, z ktorých jedna reaguje na nízku amplitúdu stimulu a druhá selektívne reaguje na vysokú amplitúdu stimulu.
Tieto výsledky majú dôležité, ale špekulatívne mechanické dôsledky: mechanický prah senzorických neurónov môže mať málo spoločného s bunkovou organizáciou mechanoreceptora, ale môže spočívať vo vlastnostiach mechanicky hradlovaných iónových kanálov.
Mechanizmy, ktoré sú základom desenzibilizácie mechanosenzitívnych katiónových prúdov v potkaních DRG neurónoch, boli nedávno odhalené.64,67 Vyplýva to z dvoch súbežných mechanizmov, ktoré ovplyvňujú vlastnosti kanálov: adaptácia a inaktivácia. Adaptácia bola prvýkrát zaznamenaná v štúdiách sluchových vláskových buniek. Operatívne ho možno opísať ako jednoduchý posun aktivačnej krivky kanála prevodníka pozdĺž osi mechanického stimulu.70-72 Adaptácia umožňuje zmyslovým receptorom zachovať si citlivosť na nové stimuly v prítomnosti existujúceho stimulu. Avšak podstatná časť mechanosenzitívnych prúdov v DRG neurónoch nemôže byť reaktivovaná po kondicionačnej mechanickej stimulácii, čo naznačuje inaktiváciu niektorých kanálov prevodníkov.64,67 Preto inaktivácia aj adaptácia pôsobia v tandeme na reguláciu mechanosenzitívnych prúdov. Tieto dva mechanizmy sú spoločné pre všetky mechanosenzitívne prúdy identifikované v potkaních DRG neurónoch, čo naznačuje, že súvisiace fyzikálno-chemické prvky určujú kinetiku týchto kanálov.64
Záverom možno povedať, že určenie vlastností endogénnych mechanosenzitívnych prúdov in vitro je rozhodujúce v snahe identifikovať transdukčné mechanizmy na molekulárnej úrovni. Variabilita pozorovaná v mechanickom prahu a adaptačná kinetika rôznych mechanicky riadených prúdov v DRG neurónoch naznačuje, že vnútorné vlastnosti iónových kanálov môžu aspoň čiastočne vysvetliť mechanický prah a kinetiku adaptácie mechanoreceptorov opísaných v desaťročiach 1960� 80 s použitím prípravkov ex vivo.
Predpokladané mechanosenzitívne proteíny
Mechanosenzitívne iónové prúdy v somatosenzorických neurónoch sú dobre charakterizované, na rozdiel od toho je málo známe o identite molekúl, ktoré sprostredkovávajú mechanotransdukciu u cicavcov. Genetické skríningy u Drosophila a C. elegans identifikovali kandidátske molekuly mechanotransdukcie, vrátane rodín TRP a degenerínových/epiteliálnych Na+ kanálov (Deg/ENaC). . Navyše, mnoho z týchto kandidátov je prítomných v kožných mechanoreceptoroch a somatosenzorických neurónoch (obr. 73).
Iónové kanály snímajúce kyselinu
ASIC patria do protónovo riadenej podskupiny degenerínových epitelových Na+ kanálov.74 Traja členovia rodiny ASIC (ASIC1, ASIC2 a ASIC3) sú exprimovaní v mechanoreceptoroch a nociceptoroch. Úloha kanálov ASIC bola skúmaná v behaviorálnych štúdiách s použitím myší s cielenou deléciou génov kanálov ASIC. Delécia ASIC1 nemení funkciu kožných mechanoreceptorov, ale zvyšuje mechanickú citlivosť aferentov inervujúcich črevo.75 Knokautované myši ASIC2 vykazujú zníženú citlivosť rýchlo sa adaptujúcich kožných LTMR. viscerálna mechano-nocicepcia aj kožná mechanosenzácia.76 Prerušenie ASIC2 znižuje mechanickú citlivosť viscerálnych aferentov a znižuje reakcie kožných HTMR na škodlivé stimuly.77
Kanál prechodného receptora
Nadrodina TRP sa u cicavcov delí na šesť podrodín.78 Takmer všetky podrodiny TRP majú členov spojených s mechanosenzáciou v rôznych bunkových systémoch.79 V senzorických neurónoch cicavcov sú však kanály TRP najlepšie známe pre snímanie tepelných informácií a sprostredkovanie neurogénneho zápalu, a iba dva kanály TRP, TRPV4 a TRPA1, sa podieľajú na odozve na dotyk. Narušenie expresie TRPV4 u myší má len mierny vplyv na akútne mechanosenzorické prahy, ale výrazne znižuje citlivosť na škodlivé mechanické stimuly.80,81 TRPV4 je kľúčovým determinantom pri formovaní odpovede nociceptívnych neurónov na osmotický stres a na mechanickú hyperalgéziu počas zápalu.82,83, Zdá sa, že TRPA1 zohráva úlohu pri mechanickej hyperalgézii. Myši s deficitom TRPA1 vykazujú precitlivenosť na bolesť. TRPA1 prispieva k transdukcii mechanických, chladových a chemických stimulov v nociceptorových senzorických neurónoch, ale zdá sa, že nie je nevyhnutný pre transdukciu vlasových buniek.84,85
Neexistuje žiadny jasný dôkaz, ktorý by naznačoval, že kanály TRP a kanály ASIC exprimované u cicavcov sú mechanicky hradené. Žiadny z týchto kanálov vyjadrený heterológne nerekapituluje elektrický podpis mechanosenzitívnych prúdov pozorovaných v ich prirodzenom prostredí. To nevylučuje možnosť, že kanály ASIC a TRP sú mechanotransduktory, vzhľadom na neistotu, či kanál mechanotransdukcie môže fungovať mimo svojho bunkového kontextu (pozri časť o SLP3).
Piezo proteíny
Coste a spolupracovníci nedávno identifikovali piezoproteíny ako sľubných kandidátov na mechanosenzorické proteíny.86,87 Stavovce majú dvoch Piezo členov, Piezo 1 a Piezo 2, predtým známe ako FAM38A a FAM38B, ktoré sú dobre zachované v mnohobunkových eukaryotoch. . Piezo 2 je v DRG hojné, zatiaľ čo Piezo 1 je sotva detegovateľné. Piezo-indukovaným mechanosenzitívnym prúdom bráni inhibícia gadolíniom, ruténiovou červeňou a GsMTx4 (toxín z tarantule Grammostola spatulata).88 Expresia Piezo 1 alebo Piezo 2 v heterológnych systémoch produkuje mechanosenzitívne prúdy, kinetika inaktivácie Piezo 2 prúdu je rýchlejšia ako Piezo 1. Podobne ako endogénne mechanosenzitívne prúdy, piezo-závislé prúdy majú reverzné potenciály okolo 0 mV a nie sú katiónovo neselektívne, pričom všetky Na+, K+, Ca2+ a Mg2+ prenikajú spodným kanálom. Podobne aj piezo-závislé prúdy sú regulované membránovým potenciálom s výrazným spomalením kinetiky prúdu pri depolarizovaných potenciáloch.86
Piezoproteíny sú nepochybne mechanosenzitívne proteíny a zdieľajú mnohé vlastnosti rýchlo sa prispôsobujúcich mechanosenzitívnych prúdov v senzorických neurónoch. Ošetrenie kultivovaných DRG neurónov s Piezo 2 krátkou interferujúcou RNA znížilo podiel neurónov s rýchlo sa adaptujúcim prúdom a znížilo percento mechanosenzitívnych neurónov.86 Transmembránové domény sú umiestnené v piezoproteínoch, ale neboli zaznamenané žiadne zjavné motívy obsahujúce póry alebo signatúry iónových kanálov. identifikované. Avšak myšací proteín Piezo 1 purifikovaný a rekonštituovaný do asymetrických lipidových dvojvrstiev a lipozómov vytvára iónové kanály citlivé na ruténiovú červeň.87 Základným krokom pri overovaní mechanotransdukcie prostredníctvom piezo kanálov je použitie in vivo prístupov na určenie funkčnej dôležitosti v dotykovej signalizácii. Informácie boli poskytnuté v Drosophila, kde delécia jedného piezočlenu znížila mechanickú odozvu na škodlivé podnety bez ovplyvnenia normálneho dotyku.89 Hoci ich štruktúru ešte treba určiť, táto nová rodina mechanosenzitívnych proteínov je sľubným predmetom budúceho výskumu, za hranicami pocitu dotyku. Napríklad nedávna štúdia na pacientoch s anémiou (hereditárna xerocytóza) ukazuje úlohu Piezo 1 pri udržiavaní homeostázy objemu erytrocytov.90
Transmembránový kanál podobný (TMC)
Nedávna štúdia naznačuje, že na mechanotransdukciu vlasových buniek sú potrebné dva proteíny, TMC1 a TMC2 Dedičná hluchota spôsobená mutáciou génu TMC91 bola hlásená u ľudí a myší.1 Prítomnosť týchto kanálov sa ešte v somatosenzorickom systéme nepreukázala , ale zdá sa, že je to dobrý nápad na preskúmanie.
Stomatin-like Protein 3 (SLP3)
Okrem transdukčných kanálov sa ukázalo, že niektoré doplnkové proteíny spojené s kanálom zohrávajú úlohu pri citlivosti na dotyk. SLP3 je exprimovaný v cicavčích DRG neurónoch. Štúdie s použitím mutantných myší, ktorým chýba SLP3, ukázali zmenu v mechanosenzácii a mechanosenzitívnych prúdoch.94,95 Presná funkcia SLP3 zostáva neznáma. Môže to byť linker medzi mechanosenzitívnym kanálom a pod ním ležiacimi mikrotubulami, ako sa navrhuje pre jeho homológ C. elegans MEC2.96 Nedávno GR. Lewinovo laboratórium navrhlo, že uväzovanie je syntetizované senzorickými neurónmi DRG a spája mechanicky citlivý iónový kanál s extracelulárnou matricou.97 Prerušením spojenia sa ruší RA-mechanosenzitívny prúd, čo naznačuje, že niektoré iónové kanály sú mechanosenzitívne iba vtedy, keď sú priviazané. RA-mechanosenzitívne prúdy sú tiež inhibované laminínom-332, matricovým proteínom produkovaným keratinocytmi, čím sa posilňuje hypotéza modulácie mechanosenzitívneho prúdu extracelulárnymi proteínmi.98
Podrodina kanálov K+
Paralelne s katiónovými depolarizačnými mechanosenzitívnymi prúdmi sa skúma prítomnosť repolarizačných mechanosenzitívnych K+ prúdov. K+ kanály v mechanosenzitívnych bunkách môžu vstúpiť do súčasnej rovnováhy a prispieť k definovaniu mechanického prahu a časového priebehu adaptácie mechanoreceptorov.
Členovia KCNK patria do rodiny dvojpórových doménových K+ kanálov (K2P). 99,100 2 K2P vykazujú pozoruhodný rozsah regulácie bunkovými, fyzikálnymi a farmakologickými činiteľmi, vrátane zmien pH, tepla, roztiahnutia a deformácie membrány. Tieto K101P sú aktívne pri pokojovom membránovom potenciáli. Niekoľko podjednotiek KCNK je exprimovaných v somatosenzorických neurónoch.2 Kanály KCNK1 (TREK-4), KCNK2 (TRAAK) a TREK-102,103 patria medzi niekoľko kanálov, pre ktoré bolo preukázané priame mechanické hradlovanie natiahnutím membrány.XNUMX
Myši s prerušeným génom KCNK2 vykazovali zvýšenú citlivosť na teplo a mierne mechanické stimuly, ale normálny prah stiahnutia na škodlivý mechanický tlak aplikovaný na zadnú labku pomocou testu Randall�Selitto. 104 Myši s deficitom KCNK2 tiež vykazujú zvýšenú tepelnú a mechanickú hyperalgéziu pri zápaloch podmienky. KCNK4 knockout myši boli precitlivené na miernu mechanickú stimuláciu a táto hypersenzitivita bola zvýšená dodatočnou inaktiváciou KCNK2.105 Zvýšená mechanosenzitivita týchto knockout myší by mohla znamenať, že natiahnutie normálne aktivuje depolarizačné aj repolarizačné mechanosenzitívne prúdy koordinovaným spôsobom, podobne ako nerovnováha depolarizujúce a repolarizujúce napäťovo riadené prúdy.
KCNK18 (TRESK) je hlavným prispievateľom k základnej vodivosti K+, ktorá reguluje pokojový membránový potenciál somatosenzorických neurónov.106 Hoci nie je známe, či je KCNK18 priamo citlivý na mechanickú stimuláciu, môže hrať úlohu pri sprostredkovaní reakcií na ľahký dotyk, ako aj bolestivé mechanické podnety. KCNK18 a v menšej miere KCNK3 sú navrhované ako molekulárny cieľ hydroxy-a-sanshool, zlúčeniny nachádzajúcej sa v zrnách korenia Schezuan, ktorá aktivuje dotykové receptory a vyvoláva u ľudí pocit mravčenia.107,108
Napäťovo závislý K+ kanál KCNQ4 (Kv7.4) je rozhodujúci pre nastavenie rýchlosti a preferencie frekvencie subpopulácie rýchlo sa adaptujúcich mechanoreceptorov u myší aj u ľudí. Mutácia KCNQ4 bola spočiatku spojená s formou dedičnej hluchoty. Je zaujímavé, že nedávna štúdia lokalizuje KCNQ4 v periférnych nervových zakončeniach kožného rýchlo sa adaptujúceho vlasového folikulu a Meissnerovho telieska. Strata funkcie KCNQ4 teda vedie k selektívnemu zvýšeniu citlivosti mechanoreceptorov na nízkofrekvenčné vibrácie. Je pozoruhodné, že ľudia s neskorým nástupom straty sluchu v dôsledku dominantných mutácií génu KCNQ4 vykazujú zvýšený výkon pri detekcii vibrácií s malou amplitúdou a nízkou frekvenciou.109
Pohľad Dr. Alexa Jimeneza
Hmat je považovaný za jeden z najzložitejších zmyslov v ľudskom tele, najmä preto, že ho nemá na starosti žiadny špecifický orgán. Namiesto toho dochádza k hmatu prostredníctvom zmyslových receptorov, známych ako mechanoreceptory, ktoré sa nachádzajú na koži a reagujú na mechanický tlak alebo deformáciu. V lysej alebo bezsrstej koži cicavcov sú štyri hlavné typy mechanoreceptorov: lamelárne telieska, hmatové telieska, Merkelové nervové zakončenia a bulbózne telieska. Mechanoreceptory fungujú tak, aby umožňovali detekciu dotyku, monitorovali polohu svalov, kostí a kĺbov, tzv. propriocepciu, a dokonca aj zvuky a pohyb tela. Pochopenie mechanizmov štruktúry a funkcie týchto mechanoreceptorov je základným prvkom pri využívaní liečebných postupov a terapií na zvládanie bolesti.
záver
Dotyk je komplexný zmysel, pretože predstavuje rôzne hmatové kvality, menovite vibrácie, tvar, textúru, potešenie a bolesť, s rôznymi rozlišovacími výkonmi. Doteraz bola korešpondencia medzi dotykovým orgánom a psychofyzickým zmyslom korelatívna a molekulárne markery špecifické pre triedu sa práve objavujú. Na uľahčenie budúcej identifikácie genomiky je teraz potrebný vývoj testov na hlodavcoch zodpovedajúcich rozmanitosti správania pri dotyku. Použitie myší, ktorým chýbajú špecifické podskupiny senzorických aferentných typov značne uľahčí identifikáciu mechanoreceptorov a senzorických aferentných vlákien spojených s konkrétnou dotykovou modalitou. Je zaujímavé, že nedávny článok otvára dôležitú otázku genetického základu mechanosenzorických vlastností u ľudí a naznačuje, že mutácia jedného génu by mohla negatívne ovplyvniť citlivosť na dotyk.110 To zdôrazňuje, že patofyziológia deficitu ľudského dotyku je z veľkej časti neznáma a určite by pokrok presnou identifikáciou podskupiny senzorických neurónov spojených s dotykovou modalitou alebo dotykovým deficitom.
Na oplátku sa dosiahol pokrok v definovaní biofyzikálnych vlastností mechano-hradlových prúdov.64 Vývoj nových techník v posledných rokoch, umožňujúcich monitorovanie zmien membránového napätia pri zaznamenávaní mechano-hradlového prúdu, sa ukázal ako cenná experimentálna metóda na opis mechanosenzitívne prúdy s rýchlou, strednou a pomalou adaptáciou (prehľad v Delmas a spolupracovníci).66,111 Budúcnosťou bude určenie úlohy súčasných vlastností v mechanizmoch adaptácie funkčne rôznorodých mechanoreceptorov a príspevku mechanosenzitívnych K+ prúdov k excitabilite LTMR a HTMR.
Molekulárna povaha mechano-gated prúdov u cicavcov je tiež perspektívnou témou výskumu do budúcnosti. Budúci výskum bude postupovať v dvoch perspektívach, najprv s cieľom určiť úlohu prídavnej molekuly, ktorá spája kanály s cytoskeletom a ktorá by bola potrebná na udelenie alebo reguláciu mechanosenzitivity iónových kanálov podobných TRP a ASIC / EnaC rodín. Po druhé, preskúmať veľkú a sľubnú oblasť príspevku piezo kanálov zodpovedaním kľúčových otázok týkajúcich sa permeačných a hradlovacích mechanizmov, podskupiny senzorických neurónov a dotykových modalít zahŕňajúcich Piezo a úlohu Piezo v neurónových bunkách spojených s mechanosenzácia.
Hmat, v porovnaní so zrakom, chuťou, zvukom a čuchom, ktoré využívajú špecifické orgány na spracovanie týchto vnemov, sa môže vyskytnúť v celom tele prostredníctvom malých receptorov známych ako mechanoreceptory. Rôzne typy mechanoreceptorov možno nájsť v rôznych vrstvách kože, kde môžu detekovať širokú škálu mechanickej stimulácie. Vyššie uvedený článok popisuje špecifické hlavné body, ktoré demonštrujú pokrok štrukturálnych a funkčných mechanizmov mechanoreceptorov spojených s hmatom. Informácie s odkazom na Národné centrum pre biotechnologické informácie (NCBI). Rozsah našich informácií je obmedzený na chiropraktiku, ako aj na poranenia a stavy chrbtice. Ak chcete prediskutovať túto tému, neváhajte sa opýtať Dr. Jimeneza alebo nás kontaktujte na adrese�915-850-0900 .
Kurátorom je Dr. Alex Jimenez
Ďalšie témy: Bolesť chrbta
Bolesti chrbta je jednou z najčastejších príčin invalidity a vymeškaných dní v práci na celom svete. Bolesti chrbta sa v skutočnosti pripisujú ako druhý najčastejší dôvod návštevy lekára, prevyšujú ju len infekcie horných dýchacích ciest. Približne 80 percent populácie aspoň raz za život zažije nejaký typ bolesti chrbta. Chrbtica je zložitá štruktúra pozostávajúca z kostí, kĺbov, väzov a svalov, okrem iného z mäkkých tkanív. Z tohto dôvodu môžu zranenia a/alebo zhoršené stavy, ako napr herné disky, môže nakoniec viesť k príznakom bolesti chrbta. Športové zranenia alebo zranenia pri automobilových nehodách sú často najčastejšou príčinou bolesti chrbta, niekedy však môžu mať aj tie najjednoduchšie pohyby bolestivé následky. Našťastie alternatívne možnosti liečby, ako je chiropraktická starostlivosť, môžu pomôcť zmierniť bolesť chrbta pomocou úprav chrbtice a manuálnych manipulácií, čo v konečnom dôsledku zlepšuje úľavu od bolesti.
1.�Moriwaki K, Yuge O. Topografické znaky kožných taktilných hypoestetických a hyperestetických abnormalít pri chronickej bolesti.�Bolesť.�1999;81:1�6. doi: 10.1016/S0304-3959(98)00257-7.�[PubMed][Cross Ref]
2.�Shim B, Kim DW, Kim BH, Nam TS, Leem JW, Chung JM. Mechanická a tepelná senzibilizácia kožných nociceptorov u potkanov s experimentálnou periférnou neuropatiou.�Neuroveda.�2005;132:193�201. doi: 10.1016/j.neuroscience.2004.12.036.�[PubMed]�[Cross Ref]
3.�Kleggetveit IP, J�rum E. Dysfunkcia veľkých a malých vlákien pri poraneniach periférnych nervov so spontánnou bolesťou alebo bez nej.�J Bolesť.�2010;11:1305�10. doi: 10.1016/j.jpain.2010.03.004.�[PubMed]�[Cross Ref]
4.�Noback ČR. Morfológia a fylogenéza vlasov.�Ann NY Acad Sci.�1951;53:476�92. doi: 10.1111/j.1749-6632.1951.tb31950.x.�[PubMed]�[Cross Ref]
9.�Brown AG, Iggo A. Kvantitatívna štúdia kožných receptorov a aferentných vlákien u mačky a králika.�J Physiol.�1967;193:707[Článok bez PMC]�[PubMed]
10.�Burgess PR, Petit D, Warren RM. Typy receptorov v mačacej chlpatej koži dodávanej myelinizovanými vláknami.�J Neurophysiol.�1968;31:833[PubMed]
11.�Driskell RR, Giangreco A, Jensen KB, Mulder KW, Watt FM. Sox2-pozitívne bunky dermálnej papily špecifikujú typ vlasového folikulu v epiderme cicavcov.�Vývoj.�2009;136:2815�23. doi: 10.1242/dev.038620.�[Článok bez PMC]�[PubMed]�[Cross Ref]
12.�Hussein MA. Celkový vzor usporiadania vlasových folikulov u potkanov a myší.�J Anat.�1971;109:307[Článok bez PMC]�[PubMed]
13.�Vielkind U, Hardy MH. Meniace sa vzory molekúl bunkovej adhézie počas vývoja vlasových folikulov z kože myší. 2. Morfogenéza folikulov vo vlasových mutantoch, Tabby a downy.�Acta Anat (Bazilej)�1996;157:183. doi: 94/10.1159.�[PubMed]�[Cross Ref]
14.�Hardy MH, Vielkind U. Meniace sa vzory molekúl bunkovej adhézie počas vývoja vlasových folikulov z kože myší. 1. Morfogenéza folikulov u myší divokého typu.�Acta Anat (Bazilej)�1996;157:169. doi: 82/10.1159.�[PubMed]�[Cross Ref]
15.�Li L, Rutlin M, Abraira VE, Cassidy C, Kus L, Gong S a kol. Funkčná organizácia kožných nízkoprahových mechanosenzorických neurónov.�Bunka.�2011;147:1615�27. doi: 10.1016/j.cell.2011.11.027.[Článok bez PMC]�[PubMed]�[Cross Ref]
16.�Brown AG, Iggo A. Kvantitatívna štúdia kožných receptorov a aferentných vlákien u mačky a králika.�J Physiol.�1967;193:707[Článok bez PMC]�[PubMed]
17.�Burgess PR, Petit D, Warren RM. Typy receptorov v mačacej chlpatej koži dodávanej myelinizovanými vláknami.�J Neurophysiol.�1968;31:833[PubMed]
18.�Vallbo A, Olausson H, Wessberg J, Norrsell U. Systém nemyelinizovaných aferentov pre neškodnú mechanorecepciu v ľudskej koži.�Brain Res.�1993;628:301�4. doi: 10.1016/0006-8993(93)90968-S.[PubMed]�[Cross Ref]
19.�Vallbo AB, Olausson H, Wessberg J. Nemyelinizované aferenty predstavujú druhý systém kódujúci hmatové stimuly ľudskej chlpatej kože.�J Neurophysiol.�1999;81:2753[PubMed]
22.�Wessberg J, Olausson H, Fernstr�m KW, Vallbo AB. Vlastnosti recepčného poľa nemyelinizovaných hmatových aferentov v ľudskej koži.�J Neurophysiol.�2003;89:1567�75. doi: 10.1152/jn.00256.2002.�[PubMed][Cross Ref]
23.�Liu Q, Vrontou S, Rice FL, Zylka MJ, Dong X, Anderson DJ. Molekulárna genetická vizualizácia vzácnej podskupiny nemyelinizovaných senzorických neurónov, ktoré dokážu rozpoznať jemný dotyk.�Nat Neurosci.�2007;10:946�8. doi: 10.1038/nn1937.�[PubMed]�[Cross Ref]
24.�Olausson H, Lamarre Y, Backlund H, Morin C, Wallin BG, Starck G a kol. Nemyelinizované hmatové aferentné signály signalizujú dotyk a premietajú sa do ostrovného kortexu.�Nat Neurosci.�2002;5: 900. doi: 4/nn10.1038.[PubMed]�[Cross Ref]
25.�Olausson H, Wessberg J, Morrison I, McGlone F, Vallbo A. Neurofyziológia nemyelinizovaných taktilných aferentov.�Neurosci Biobehav Rev.�2010;34:185. doi: 91/j.neubiorev.10.1016.�[Recenzia][PubMed]�[Cross Ref]
26.�Kr�mer HH, Lundblad L, Birklein F, Linde M, Karlsson T, Elam M a kol. Aktivácia kortikálnej siete bolesti mäkkou taktilnou stimuláciou po injekcii sumatriptánu.�Bolesť.�2007;133:72. doi: 8/j.pain.10.1016.�[PubMed]�[Cross Ref]
27.�Applebaum AE, Beall JE, Foreman RD, Willis WD. Organizácia a receptívne polia neurónov spinotalamického traktu primátov.�J Neurophysiol.�1975;38:572[PubMed]
28.�White JC, Sweet WH. Účinnosť chordotómie pri fantómovej bolesti po amputácii.�Psychiatria AMA Arch Neurol.�1952;67:315[PubMed]
29.�Halata Z, Grim M, Bauman KI. Friedrich Sigmund Merkel a jeho „Merkelova bunka“, morfológia, vývoj a fyziológia: prehľad a nové výsledky.�Anat Rec A Discov Mol Cell Evol Biol.�2003;271:225. doi: 39/ar.a.10.1002.�[PubMed]�[Cross Ref]
30.�Morrison KM, Miesegaes GR, Lumpkin EA, Maricich SM. Cicavčie Merkelove bunky pochádzajú z epidermálnej línie.�Dev Biol.�2009;336:76. doi: 83/j.ydbio.10.1016.[Článok bez PMC]�[PubMed]�[Cross Ref]
31.�Van Keymeulen A, Mascre G, Youseff KK, Harel I, Michaux C, De Geest N a kol. Epidermálne progenitory spôsobujú vznik Merkelových buniek počas embryonálneho vývoja a homeostázy dospelých.�J Cell Biol.�2009;187: 91 � 100. doi: 10.1083/jcb.200907080.�[Článok bez PMC]�[PubMed]�[Cross Ref]
32.�Ebara S, Kumamoto K, Baumann KI, Halata Z. Trojrozmerné analýzy dotykových kupol vo chlpatej koži mačacej labky odhaľujú morfologické substráty pre komplexné senzorické spracovanie.�Neurosci Res.�2008;61:159. doi: 71/j.neures.10.1016.�[PubMed]�[Cross Ref]
33.�Guinard D, Usson Y, Guillermet C, Saxod R. Merkel komplexy ľudskej digitálnej kože: trojrozmerné zobrazovanie pomocou konfokálnej laserovej mikroskopie a dvojitej imunofluorescencie.�J Comp Neurol.�1998;398:98�104. doi: 10.1002/(SICI)1096-9861(19980817)398:1<98::AID-CNE6>3.0.CO;2-4.�[PubMed][Cross Ref]
34.�Reinisch CM, Tschachler E. Dotyková kupola v ľudskej koži je zásobovaná rôznymi typmi nervových vlákien.�Ann Neurol.�2005;58:88. doi: 95/ana.10.1002.�[PubMed]�[Cross Ref]
35.�Maricich SM, Morrison KM, Mathes EL, Brewer BM. Hlodavce sa pri úlohách rozlišovania textúr spoliehajú na Merkelove bunky.�J Neurosci.�2012;32:3296�300. doi: 10.1523/JNEUROSCI.5307-11.2012.[Článok bez PMC]�[PubMed]�[Cross Ref]
36.�Ikeda I, Yamashita Y, Ono T, Ogawa H. Selektívna fototoxická deštrukcia potkaních Merkelových buniek ruší reakcie pomaly sa adaptujúcich mechanoreceptorových jednotiek typu I.�J Physiol.�1994;479:247[Článok bez PMC][PubMed]
37.�Maricich SM, Wellnitz SA, Nelson AM, Lesniak DR, Gerling GJ, Lumpkin EA a kol. Merkelove bunky sú nevyhnutné pre ľahké reakcie.�Veda.�2009;324:1580�2. doi: 10.1126/science.1172890.[Článok bez PMC]�[PubMed]�[Cross Ref]
38.�Diamond J, Holmes M, sestra CA. Sú recipročné synapsie Merkelovej bunky a neuritov zahrnuté v iniciácii hmatových reakcií v koži salamandra?�J Physiol.�1986;376:101[Článok bez PMC][PubMed]
39.�Yamashita Y, Akaike N, Wakamori M, Ikeda I, Ogawa H. Prúdy závislé od napätia v izolovaných jednotlivých Merkelových bunkách potkanov.�J Physiol.�1992;450:143[Článok bez PMC]�[PubMed]
40.�Wellnitz SA, Lesniak DR, Gerling GJ, Lumpkin EA. Pravidelnosť nepretržitého odpaľovania odhaľuje dve populácie pomaly sa prispôsobujúcich dotykových receptorov v chlpatej koži myší.�J Neurophysiol.�2010;103:3378�88. doi: 10.1152/jn.00810.2009.�[Článok bez PMC]�[PubMed]�[Cross Ref]
41.�Nandasena BG, Suzuki A, Aita M, Kawano Y, Nozawa-Inoue K, Maeda T. Imunolokalizácia aquaporínu-1 v mechanoreceptívnych Ruffiniho zakončeniach v periodontálnom väze.�Brain Res.�2007;1157:32-40. doi: 10.1016/j.brainres.2007.04.033.�[PubMed]�[Cross Ref]
42.�Rahman F, Harada F, Saito I, Suzuki A, Kawano Y, Izumi K a kol. Detekcia acid-sensing ion channel 3 (ASIC3) v periodontálnych Ruffiniho zakončeniach myších rezákov.�Neurosci Lett.�2011;488:173. doi: 7/j.neulet.10.1016.�[PubMed]�[Cross Ref]
43.�Johnson KO. Úlohy a funkcie kožných mechanoreceptorov.�Curr Opin Neurobiol.�2001;11:455�61. doi: 10.1016/S0959-4388(00)00234-8.�[Review]�[PubMed]�[Cross Ref]
44.�Wende H, Lechner SG, Cheret C, Bourane S, Kolanczyk ME, Pattyn A a kol. Transkripčný faktor c-Maf riadi vývoj a funkciu dotykového receptora.�Veda.�2012;335:1373�6. doi: 10.1126/science.1214314.�[PubMed]�[Cross Ref]
45.�Mendelson M, Lowenstein WR. Mechanizmy adaptácie receptorov.�Veda.�1964;144:554�5. doi: 10.1126/science.144.3618.554.�[PubMed]�[Cross Ref]
46.�Loewenstein WR, Mendelson M. Komponenty adaptácie receptora v paciniovom teliesku.�J Physiol.�1965;177:377[Článok bez PMC]�[PubMed]
47.�Pawson L, Prestia LT, Mahoney GK, G��l� B, Cox PJ, Pack AK. GABAergická/glutamátergická-gliová/neurónová interakcia prispieva k rýchlej adaptácii v paciniových telieskach.�J Neurosci.�2009;29:2695�705. doi: 10.1523/JNEUROSCI.5974-08.2009.�[Článok bez PMC]�[PubMed]�[Cross Ref]
48.�Basbaum AI, Jessell TM. Vnímanie bolesti. In: Kandel ER, Schwartz JH, Jessell TM, eds. Princípy neurálnej vedy. Štvrté vydanie. The McGraw-Hill Compagies, 2000: 472-490.
49.�Bourane S, Garces A, Venteo S, Pattyn A, Hubert T, Fichard A a kol. Podtypy nízkoprahových mechanoreceptorov selektívne exprimujú MafA a sú špecifikované signalizáciou Ret.�Neurón.�2009;64:857. doi: 70/j.neuron.10.1016.�[PubMed]�[Cross Ref]
50.�Kramer I, Sigrist M, de Nooij JC, Taniuchi I, Jessell TM, Arber S. Úloha signalizácie transkripčného faktora Runx pri diverzifikácii senzorických neurónov ganglií dorzálnych koreňov.�Neurón.�2006;49:379. doi: 93/j.neuron.10.1016.�[PubMed]�[Cross Ref]
51.�Luo W, Enomoto H, Rice FL, Milbrandt J, Ginty DD. Molekulárna identifikácia rýchlo sa adaptujúcich mechanoreceptorov a ich vývojová závislosť od ret signalizácie.�Neurón.�2009;64:841. doi: 56/j.neuron.10.1016.�[Článok bez PMC]�[PubMed]�[Cross Ref]
52.�Vallbo AB, Hagbarth KE. Aktivita mechanoreceptorov kože zaznamenaná perkutánne u bdelých ľudských subjektov.�Exp Neurol.�1968;21:270�89. doi: 10.1016/0014-4886(68)90041-1.�[PubMed]�[Cross Ref]
53.�Macefield VG. Fyziologické charakteristiky nízkoprahových mechanoreceptorov v kĺboch, svaloch a koži u ľudských subjektov.�Clin Exp Pharmacol Physiol.�2005;32:135�44. doi: 10.1111/j.1440-1681.2005.04143.x.�[Review]�[PubMed]�[Cross Ref]
54.�Koizumi S, Fujishita K, Inoue K, Shigemoto-Mogami Y, Tsuda M, Inoue K. Ca2+ vlny v keratinocytoch sa prenášajú do senzorických neurónov: zapojenie extracelulárnej aktivácie ATP a P2Y2 receptora.�Biochem J.�2004;380:329-38. doi: 10.1042/BJ20031089.�[Článok bez PMC]�[PubMed][Cross Ref]
55.�Azorin N, Raoux M, Rodat-Despoix L, Merrot T, Delmas P, Crest M. Signalizácia ATP je rozhodujúca pre odpoveď ľudských keratinocytov na mechanickú stimuláciu hypoosmotickým šokom.�Exp Dermatol.�2011;20:401�7. doi: 10.1111/j.1600-0625.2010.01219.x.�[PubMed]�[Cross Ref]
56.�Amano M, Fukata Y, Kaibuchi K. Regulácia a funkcie Rho-asociovanej kinázy.�Exp Cell Res.�2000;261:44�51. doi: 10.1006/excr.2000.5046.�[Recenzia]�[PubMed]�[Cross Ref]
57.�Koyama T, Oike M, Ito Y. Zapojenie Rho-kinázy a tyrozínkinázy do uvoľňovania ATP indukovaného hypotonickým stresom v endotelových bunkách bovinnej aorty.�J Physiol.�2001;532:759�69. doi: 10.1111/j.1469-7793.2001.0759e.x.�[Článok bez PMC]�[PubMed]�[Cross Ref]
60.�Drew LJ, Wood JN, Cesare P. Výrazné mechanosenzitívne vlastnosti senzorických neurónov citlivých a necitlivých na kapsaicín.�J Neurosci.�2002;22:RC228.�[PubMed]
61.�Drew LJ, Rohrer DK, Price MP, Blaver KE, Cockayne DA, Cesare P a kol. Iónové kanály ASIC2 a ASIC3 citlivé na kyselinu neprispievajú k mechanicky aktivovaným prúdom v senzorických neurónoch cicavcov.�J Physiol.�2004;556:691�710. doi: 10.1113/jphysiol.2003.058693.�[Článok bez PMC]�[PubMed]�[Cross Ref]
62.�McCarter GC, Levine JD. Iónový základ mechanotransdukčného prúdu v neurónoch dorzálnych koreňových ganglií dospelých potkanov.�Mol Pain.�2006;2:28. doi: 10.1186/1744-8069-2-28.�[Článok bez PMC]�[PubMed]�[Cross Ref]
63.�Coste B, Crest M, Delmas P. Farmakologická disekcia a distribúcia NaN/Nav1.9, Ca2+ prúdov typu T a mechanicky aktivovaných katiónových prúdov v rôznych populáciách neurónov DRG.�J Gen Physiol.�2007;129:57. doi: 77/jgp.10.1085.�[Článok bez PMC]�[PubMed]�[Cross Ref]
65.�Drew LJ, Wood JN. FM1-43 je permanentný blokátor mechanosenzitívnych iónových kanálov v senzorických neurónoch a inhibuje behaviorálne reakcie na mechanické stimuly.�Mol Pain.�2007;3:1. doi: 10.1186/1744-8069-3-1.�[Článok bez PMC]�[PubMed]�[Cross Ref]
66.�Hao J, Delmas P. Záznam mechanosenzitívnych prúdov pomocou piezoelektricky poháňaného mechanostimulátora.�Nat Protoc.�2011;6:979. doi: 90/nprot.10.1038.�[PubMed]�[Cross Ref]
67.�Rugiero F, Drew LJ, Wood JN. Kinetické vlastnosti mechanicky aktivovaných prúdov v miechových senzorických neurónoch.�J Physiol.�2010;588:301�14. doi: 10.1113/jphysiol.2009.182360.�[Článok bez PMC]�[PubMed][Cross Ref]
68.�Hu J, Lewin GR. Mechanosenzitívne prúdy v neuritoch kultivovaných myších senzorických neurónov.�J Physiol.�2006;577:815�28. doi: 10.1113/jphysiol.2006.117648.�[Článok bez PMC]�[PubMed]�[Cross Ref]
69.�Bhattacharya MR, Bautista DM, Wu K, Haeberle H, Lumpkin EA, Julius D. Radiálny úsek odhaľuje odlišné populácie mechanosenzitívnych somatosenzorických neurónov cicavcov.�Proc Natl Acad Sci USA A.�2008;105:20015�20. doi: 10.1073/pnas.0810801105.�[Článok bez PMC]�[PubMed]�[Cross Ref]
70.�Crawford AC, Evans MG, Fettiplace R. Aktivácia a adaptácia transduktorových prúdov vo vlasových bunkách korytnačiek.�J Physiol.�1989;419:405[Článok bez PMC]�[PubMed]
71.�Ricci AJ, Wu YC, Fettiplace R. Endogénny vápnikový pufor a časový priebeh adaptácie prevodníka v sluchových vláskových bunkách.�J Neurosci.�1998;18:8261[PubMed]
72.�Vollrath MA, Kwan KY, Corey DP. Mikrostroje mechanotransdukcie vo vlasových bunkách.�Annu Rev Neurosci.�2007;30:339. doi: 65/annurev.neuro.10.1146.�[Recenzia]�[Článok bez PMC]�[PubMed]�[Cross Ref]
74.�Waldmann R, Lazdunski MH. H(+)-hradené katiónové kanály: senzory neurónových kyselín v rodine iónových kanálov NaC/DEG.�Curr Opin Neurobiol.�1998;8:418�24. doi: 10.1016/S0959-4388(98)80070-6.[Review]�[PubMed]�[Cross Ref]
75.�Page AJ, Brierley SM, Martin CM, Martinez-Salgado C, Wemmie JA, Brennan TJ, et al. Iónový kanál ASIC1 prispieva k viscerálnej, ale nie kožnej funkcii mechanoreceptorov.�Gastroenterológia.�2004;127:1739�47. doi: 10.1053/j.gastro.2004.08.061.�[PubMed]�[Cross Ref]
76.�Price MP, McIlwrath SL, Xie J, Cheng C, Qiao J, Tarr DE a kol. Katiónový kanál DRASIC prispieva k detekcii kožného dotyku a kyslých stimulov u myší.�Neurón.�2001;32:1071�83. doi: 10.1016/S0896-6273(01)00547-5.�[Erratum in: Neuron 2002 Jul 18;35] [2]�[PubMed]�[Cross Ref]
77.�Roza C, Puel JL, Kress M, Baron A, Diochot S, Lazdunski M, et al. Knockout kanálu ASIC2 u myší nezhoršuje kožnú mechanosenzáciu, viscerálnu mechanonocicepciu a sluch.�J Physiol.�2004;558:659�69. doi: 10.1113/jphysiol.2004.066001.�[Článok bez PMC]�[PubMed]�[Cross Ref]
78.�Damann N, Voets T, Nilius B. TRP v našich zmysloch.�Curr Biol.�2008;18:R880. doi: 9/j.cub.10.1016.�[Recenzia]�[PubMed]�[Cross Ref]
79.�Christensen AP, Corey DP. TRP kanály v mechanosenzácii: priama alebo nepriama aktivácia?�Nat Rev Neurosci.�2007;8:510�21. doi: 10.1038/nrn2149.�[Recenzia]�[PubMed]�[Cross Ref]
80.�Liedtke W, Tobin DM, Bargmann CI, Friedman JM. Cicavčí TRPV4 (VR-OAC) riadi behaviorálne reakcie na osmotické a mechanické stimuly u Caenorhabditis elegans.�Proc Natl Acad Sci USA A.�2003;100(Suppl 2):14531�6. doi: 10.1073/pnas.2235619100.�[Článok bez PMC]�[PubMed]�[Cross Ref]
81.�Suzuki M, Mizuno A, Kodaira K, Imai M. Zhoršený pocit tlaku u myší bez TRPV4.�J Biol Chem.�2003;278:22664�8. doi: 10.1074/jbc.M302561200.�[PubMed]�[Cross Ref]
82.�Liedtke W, Choe Y, Mart�-Renom MA, Bell AM, Denis CS, Sali A a kol. Osmoticky aktivovaný kanál súvisiaci s vaniloidným receptorom (VR-OAC), kandidátsky osmoreceptor u stavovcov.�Bunka.�2000;103:525�35. doi: 10.1016/S0092-8674(00)00143-4.�[Článok bez PMC]�[PubMed]�[Cross Ref]
83.�Alessandri-Haber N, Dina OA, Yeh JJ, Parada CA, Reichling DB, Levine JD. Prechodný receptorový potenciál vaniloidu 4 je nevyhnutný pri neuropatickej bolesti u potkanov vyvolanej chemoterapiou.�J Neurosci.�2004;24:4444�52. doi: 10.1523/JNEUROSCI.0242-04.2004.�[Erratum in: J Neurosci. jún 2004;24] [23][PubMed]�[Cross Ref]
84.�Bautista DM, Jordt SE, Nikai T, Tsuruda PR, Read AJ, Poblete J a kol. TRPA1 sprostredkúva zápalové účinky environmentálnych dráždivých látok a proalgetických činidiel.�Bunka.�2006;124:1269�82. doi: 10.1016/j.cell.2006.02.023.�[PubMed]�[Cross Ref]
85.�Kwan KY, Allchorne AJ, Vollrath MA, Christensen AP, Zhang DS, Woolf CJ a kol. TRPA1 prispieva k studenej, mechanickej a chemickej nocicepcii, ale nie je nevyhnutný pre transdukciu vlasových buniek.�Neurón.�2006;50:277. doi: 89/j.neuron.10.1016.�[PubMed]�[Cross Ref]
86.�Coste B, Mathur J, Schmidt M, Earley TJ, Ranade S, Petrus MJ a kol. Piezo1 a Piezo2 sú základnými komponentmi odlišných mechanicky aktivovaných katiónových kanálov.�Veda.�2010;330:55�60. doi: 10.1126/science.1193270.�[Článok bez PMC]�[PubMed]�[Cross Ref]
87.�Coste B, Xiao B, Santos JS, Syeda R, Grandl J, Spencer KS a kol. Piezoproteíny sú pórotvorné podjednotky mechanicky aktivovaných kanálov.�Príroda.�2012;483:176�81. doi: 10.1038/príroda10812.[Článok bez PMC]�[PubMed]�[Cross Ref]
88.�Bae C, Sachs F, Gottlieb PA. Mechanosenzitívny iónový kanál Piezo1 je inhibovaný peptidom GsMTx4.�Biochémia.�2011;50:6295�300. doi: 10.1021/bi200770q.�[Článok bez PMC]�[PubMed][Cross Ref]
89.�Kim SE, Coste B, Chadha A, Cook B, Patapoutian A. Úloha Drosophila Piezo v mechanickej nocicepcii.�Príroda.�2012;483:209�12. doi: 10.1038/nature10801.�[Článok bez PMC]�[PubMed][Cross Ref]
90.�Zarychanski R, Schulz VP, Houston BL, Maksimova Y, Houston DS, Smith B a kol. Mutácie v mechanotransdukčnom proteíne PIEZO1 sú spojené s dedičnou xerocytózou.�Krv.�2012;120:1908�15. doi: 10.1182/blood-2012-04-422253.�[Článok bez PMC]�[PubMed]�[Cross Ref]
91.�Kawashima Y, G�l�oc GS, Kurima K, Labay V, Lelli A, Asai Y a kol. Mechanotransdukcia vo vláskových bunkách vnútorného ucha myší vyžaduje gény podobné transmembránovým kanálom.�J Clin Invest.�2011;121:4796�809. doi: 10.1172/JCI60405.�[Článok bez PMC]�[PubMed]�[Cross Ref]
92.�Tlili A, Rebeh IB, Aifa-Hmani M, Dhouib H, Moalla J, Tlili-Chouchne J, et al. TMC1, ale nie TMC2, je zodpovedný za autozomálne recesívne nesyndromové poškodenie sluchu v tuniských rodinách.�Audiol Neurootol.�2008;13:213. doi: 8/10.1159.�[PubMed]�[Cross Ref]
93.�Manji SS, Miller KA, Williams LH, Dahl HH. Identifikácia troch nových kmeňov myší so stratou sluchu s mutáciami v géne Tmc1.�Am J Pathol.�2012;180:1560�9. doi: 10.1016/j.ajpath.2011.12.034.[PubMed]�[Cross Ref]
94.�Wetzel C, Hu J, Riethmacher D, Benckendorff A, Harder L, Eilers A a kol. Proteín stomatínovej domény nevyhnutný pre pocit dotyku u myši.�Príroda.�2007;445:206�9. doi: 10.1038/nature05394.�[PubMed][Cross Ref]
95.�Martinez-Salgado C, Benckendorff AG, Chiang LY, Wang R, Milenkovic N, Wetzel C, a kol. Mechanotransdukcia stomatínu a senzorických neurónov.�J Neurophysiol.�2007;98:3802�8. doi: 10.1152/jn.00860.2007.[PubMed]�[Cross Ref]
96.�Huang M, Gu G, Ferguson EL, Chalfie M. Proteín podobný stomatínu potrebný na mechanosenzáciu u C. elegans.�Príroda.�1995;378:292�5. doi: 10.1038/378292a0.�[PubMed]�[Cross Ref]
97.�Hu J, Chiang LY, Koch M, Lewin GR. Dôkaz o proteínovom reťazci zapojenom do somatického dotyku.�EMBO J.�2010;29:855. doi: 67/emboj.10.1038.�[Článok bez PMC]�[PubMed]�[Cross Ref]
98.�Chiang LY, Poole K, Oliveira BE, Duarte N, Sierra YA, Bruckner-Tuderman L a kol. Laminín-332 koordinuje mechanotransdukciu a bifurkáciu rastového kužeľa v senzorických neurónoch.�Nat Neurosci.�2011;14: 993 � 1000 10.1038. doi: 2873/nn.XNUMX.�[PubMed]�[Cross Ref]
99.�Lesage F, Guillemare E, Fink M, Duprat F, Lazdunski M, Romey G a kol. TWIK-1, všadeprítomný ľudský slabo vnútorne usmerňujúci K+ kanál s novou štruktúrou.�EMBO J.�1996;15:1004.[Článok bez PMC]�[PubMed]
100.�Lesage F. Farmakológia draslíkových kanálov neurónového pozadia.�Neurofarmakológia.�2003;44:1�7. doi: 10.1016/S0028-3908(02)00339-8.�[Review]�[PubMed]�[Cross Ref]
101.�Medhurst AD, Rennie G, Chapman CG, Meadows H, Duckworth MD, Kelsell RE a kol. Distribučná analýza ľudských dvoch pórových domén draslíkových kanálov v tkanivách centrálneho nervového systému a periférie.�Brain Res Mol Brain Res.�2001;86:101�14. doi: 10.1016/S0169-328X(00)00263-1.�[PubMed][Cross Ref]
102.�Mainingret F, Patel AJ, Lesage F, Lazdunski M, Honor� E. Mechano- alebo kyslá stimulácia, dva interaktívne režimy aktivácie draslíkového kanála TREK-1.�J Biol Chem.�1999;274:26691�6. doi: 10.1074/jbc.274.38.26691.�[PubMed]�[Cross Ref]
103.�Maingret F, Fosset M, Lesage F, Lazdunski M, Honor� E. TRAAK je cicavčí neurónový mechanicky riadený K+ kanál.�J Biol Chem.�1999;274:1381�7. doi: 10.1074/jbc.274.3.1381.�[PubMed][Cross Ref]
104.�Alloui A, Zimmermann K, Mamet J, Duprat F, No�l J, Chemin J a kol. TREK-1, K+ kanál zapojený do polymodálneho vnímania bolesti.�EMBO J.�2006;25:2368-76. doi: 10.1038/sj.emboj.7601116.[Článok bez PMC]�[PubMed]�[Cross Ref]
105.�No�l J, Zimmermann K, Busserolles J, Deval E, Alloui A, Diochot S, a kol. Mechanicky aktivované K+ kanály TRAAK a TREK-1 ovládajú vnímanie tepla aj chladu.�EMBO J.�2009;28:1308. doi: 18/emboj.10.1038.�[Článok bez PMC]�[PubMed]�[Cross Ref]
106.�Dobler T, Springauf A, Tovornik S, Weber M, Schmitt A, Sedlmeier R, et al. TRESK dvojpórové K+ kanály tvoria významnú zložku draslíkových prúdov na pozadí v myšacích neurónoch dorzálnych koreňových ganglií.�J Physiol.�2007;585:867. doi: 79/jphysiol.10.1113.[Článok bez PMC]�[PubMed]�[Cross Ref]
107.�Bautista DM, Sigal YM, Milstein AD, Garrison JL, Zorn JA, Tsuruda PR a kol. Štipľavé látky zo sečuánskych paprík vzrušujú senzorické neuróny inhibíciou dvojpórových draslíkových kanálov.�Nat Neurosci.�2008;11: 772 � 9 10.1038. doi: 2143/nn.XNUMX.�[Článok bez PMC]�[PubMed]�[Cross Ref]
108.�Lennertz RC, Tsunozaki M, Bautista DM, Stucky CL. Fyziologický základ brnenia parestézie vyvolanej hydroxy-alfa-sanshool.�J Neurosci.�2010;30:4353�61. doi: 10.1523/JNEUROSCI.4666-09.2010.�[Článok bez PMC]�[PubMed]�[Cross Ref]
109.�Heidenreich M, Lechner SG, Vardanyan V, Wetzel C, Cremers CW, De Leenheer EM a kol. KCNQ4 K(+) kanály vylaďujú mechanoreceptory pre normálny dotykový pocit u myši a človeka.�Nat Neurosci.�2012;15: 138 � 45 10.1038. doi: 2985/nn.XNUMX.�[PubMed]�[Cross Ref]
110.�Frenzel H, Bohlender J, Pinsker K, Wohlleben B, Tank J, Lechner SG a kol. Genetický základ pre mechanosenzorické znaky u ľudí.�PLoS Biol.�2012;10:e1001318. doi: 10.1371/journal.pbio.1001318.[Článok bez PMC]�[PubMed]�[Cross Ref]
111.�Delmas P, Hao J, Rodat-Despoix L. Molekulárne mechanizmy mechanotransdukcie v senzorických neurónoch cicavcov.�Nat Rev Neurosci.�2011;12:139�53. doi: 10.1038/nrn2993.�[PubMed]�[Cross Ref]
Mechanoreceptívna bolesť: Podľa CDC, �viac ako 50 % dospelých v USA (125 miliónov) malo muskuloskeletálny porucha bolesti v roku 2012.�
�V roku 40 viac ako 2012 % dospelých s poruchou muskuloskeletálnej bolesti z akéhokoľvek dôvodu použilo doplnkový zdravotný prístup. To bolo výrazne vyššie ako u osôb bez poruchy muskuloskeletálnej bolesti (24.1 %). Používanie doplnkových zdravotných prístupov z akéhokoľvek dôvodu medzi osobami s bolesťami krku alebo problémami bolo viac ako dvakrát vyššie ako u osôb bez týchto problémov.�
�Medzi dospelými s poruchou muskuloskeletálnej bolesti bolo použitie akéhokoľvek doplnkového zdravotného prístupu najvyššie medzi osobami s bolesťou krku alebo problémami (50.6 %), po ktorých nasledovali osoby s inými muskuloskeletálnymi problémami (46.2 %).
�Používanie doplnkových zdravotných prístupov z akéhokoľvek dôvodu medzi osobami s bolesťami krku alebo problémami bolo viac ako dvojnásobne vyššie ako u osôb bez týchto problémov.�
Mechanoreceptory sú senzorické receptory, ktoré reagujú na mechanický tlak alebo skreslenie.
Patria sem kožné receptory pre dotyk, receptory monitorujúce dĺžku a napätie svalov, sluchové a vestibulárne receptory a iné.
TEÓRIA RIADENIA BRÁNY BOLESTI
Bezbolestný vstup zatvára brány bolestivému vstupu.
To zabraňuje tomu, aby sa pocity bolesti dostali do vyšších kortikálnych úrovní
Malý priemer aferentov (bolesť) bráni inhibícii bolesti
Veľký priemer aferentov (vibrácie) má tendenciu vyvolávať inhibíciu bolesti.
Táto teória tvrdí, že nenociceptívne vlákna môžu interferovať so signálmi z vlákien bolesti, a preto inhibujú bolesť.
Veľký priemer A? vlákna sú nonnociceptívne (neprenášajú bolestivé podnety) a inhibujú účinky streľby A? a C vlákna.
CHRBTOVÝ STĹPEC MEDIÁLNA LEMNISKÁLNA DRÁHA
VYUŽITIE PERIFERNÝCH MECHANORECEPTOROV NA ZMENU VNÍMANIA BOLESTI
Ako môžeme pomôcť?
ODBEROVÝ REFLEX
Excitovaný aferentný neurón stimuluje excitačné interneuróny, ktoré následne stimulujú eferentné motorické neuróny zásobujúce biceps, sval na paži, ktorý ohýba (ohýba) lakťový kĺb. Stiahnutie bicepsu odtiahne ruku od horúceho sporáka.
Aferentný neurón tiež stimuluje inhibičné interneuróny, ktoré následne inhibujú eferentné neuróny zásobujúce triceps, aby zabránili jeho kontrakcii. Tento typ neuronálneho spojenia zahŕňajúci stimuláciu nervového zásobovania jedného svalu a súčasnú inhibíciu nervov jeho antagonistického svalu je známy ako recipročná inhibícia.
Aferentný neurón stále stimuluje ďalšie interneuróny, ktoré prenášajú signál hore miechou do mozgu prostredníctvom vzostupnej dráhy. Až keď impulz dosiahne senzorickú oblasť kôry, človek si uvedomí bolesť, jej lokalizáciu a typ stimulu. Navyše, keď impulz zasiahne mozog, informácie sa môžu uložiť ako pamäť a človek môže premýšľať o tom, čo sa stalo.
RECEPTOROVÁ TERAPIA
Úpravy
Aktivácia mechanoreceptorov kĺbov prostredníctvom chiropraktických úprav môže modulovať a zatieniť vnímanie vlákien s menším priemerom v mozgu.
Opakovanie aktivácie kĺbových mechanoreceptorov môže vytvoriť pozitívnu plasticitu v aferentných dráhach.
Pozitívna plasticita môže zastaviť bolesť
chvenie
Vibračná stimulácia pri špecifických frekvenciách môže zmeniť vnímanie bolesti
Opakovanie aktivácie Merkelových diskov a Meissnerových teliesok môže vytvoriť pozitívnu plasticitu v aferentných dráhach.
Opäť platí, že pozitívna plasticita môže zastaviť bolesť
VIBRÁCIE
�Tento typ zariadenia využíva sínusové vibrácie a ponúka nepretržitú voliteľnú amplitúdu 0-5.2 mm v závislosti od polohy nohy a voliteľnú frekvenciu 5-30 Hz.�
�Tréning WBV sa javí ako účinný, bezpečný a vhodný zásah pre pracujúcich zamestnancov v sede s chronickou bolesťou krížov.�
Homotopická vibro-taktilná stimulácia viedla k 40% zníženiu tepelnej bolesti vo všetkých skupinách subjektov. Zdá sa, že rozptýlenie neovplyvňuje experimentálne hodnotenie bolesti.�
�Vibrotaktilná stimulácia účinne aktivovala analgetické mechanizmy nielen u NC, ale aj u pacientov s chronickou muskuloskeletálnou bolesťou, vrátane FM.�
ĽAHKÝ DOTYK
�Celkovo 44 zdravých dobrovoľníkov pociťovalo tepelnú bolesť a CT optimálne (pomalé čistenie) a CT suboptimálne (rýchle čistenie alebo vibrácie) stimuly. Boli použité tri rôzne experimentálne paradigmy: Súbežná aplikácia tepelnej bolesti a hmatovej (pomalé čistenie alebo vibrácie) stimulácie; Pomalé kefovanie, aplikované na rôzne trvanie a intervaly, pred tepelnou bolesťou; Pomalé verzus rýchle čistenie zubov pred tepelnou bolesťou.�
U ľudí patria hlavné oblasti mozgu, ktoré prijímajú informácie C-LTMR, do somatosenzorického systému a ovplyvňujú spracovávacie mozgové siete, ako je kontralaterálny zadný ostrovný kortex alebo mediálny prefrontálny kortex. Intenzita CT cieleného dotyku je kódovaná v primárnej a sekundárnej somatosenzorickej kôre (S1 kontralaterálna, S2 bilaterálna), zatiaľ čo príjemnosť je kódovaná v prednej cingulárnej kôre. C-LTMR tiež aktivujú oblasti zapojené do spracovania odmien (putamen a orbitofrontálny kortex) a do spracovania sociálnych stimulov (posterior superior temporal sulcus).
VŠETKO PERIFÉRNE MÁ CENTRÁLNY NÁSLEDOK
CASE STUDY
47-ročný muž utrpel ľavú CMP v októbri 2017.
Od nehody nepohol pravou stranou tela.
Prezentovaný na našej klinike, pretože sa k tomu chce „vrátiť“.
NAJLEPŠIE FYZICKÉ VYŠETRENIE
Dyzartria
Zmenené vnímanie bolesti
Ťažkosti s jednoduchou matematikou
Flaccid na RUE a RLE
NAJLEPŠIE FYZICKÉ VYŠETRENIE
Pacient nemal žiadny pohyb, kým sme nezačali testovať pocity a reflexy:
ALLODYNIA:�Týka sa centrálnej senzibilizácie bolesti (zvýšená odozva neurónov) po normálne nebolestivej, často sa opakujúcej stimulácii.
Alodýnia môže viesť k spusteniu reakcie na bolesť zo stimulov, ktoré zvyčajne nevyvolávajú bolesť.
Teplotné alebo fyzické podnety môžu vyvolať alodýniu, ktorá môže mať pocit pálenia a často sa vyskytuje po poranení miesta.
Alldynia sa líši od hyperalgézie, extrémnej, prehnanej reakcie na podnet, ktorá je normálne bolestivá.
Prečo lokalizované poškodenie alebo zranenie spôsobené traumou vedie u niektorých pacientov k chronickej, nezvládnuteľnej bolesti? Čo má na starosti premenu lokálneho poranenia s akútnou bolesťou na chronický bolestivý stav? Prečo niektoré bolesti reagujú na protizápalové lieky a/alebo lieky, zatiaľ čo iné formy bolesti vyžadujú opiáty?
Bolesť je zložitý proces zahŕňajúci periférny nervový systém (PNS) aj centrálny nervový systém (CNS). Poranenie tkaniva spúšťa PNS, ktorý prenáša signály cez miechu do mozgu, v ktorom dochádza k vnímaniu bolesti. Čo však spôsobuje, že sa intenzívny zážitok bolesti rozvinie do neutíchajúceho javu? Dá sa tomu nejako zabrániť? Dôkazy tomu nasvedčujú chronická bolesť je výsledkom kombinácie mechanizmov, ako sú neurologické „spomienky“ predchádzajúcej bolesti.
Nociception: Najjednoduchšia cesta
Akútna alebo nociceptívna bolesť je charakterizovaná ako pravidelná skúsenosť s nepríjemným pocitom, ktorý sa vyskytuje v reakcii na veľmi základné poškodenie alebo zranenie. Je ochranný, varuje nás, aby sme sa vzdialili od pôvodu urážky a postarali sa o traumu. Mechanizmy, ktoré vytvárajú nociceptívnu bolesť, zahŕňajú transdukciu, ktorá rozširuje vonkajšiu traumatickú stimuláciu na elektrickú aktivitu v špecializovaných nociceptívnych primárnych aferentných nervoch. Aferentné nervy potom vedú senzorické informácie z PNS do CNS.
V CNS sú údaje o bolesti prenášané primárnymi senzorickými neurónmi do centrálnych projekčných buniek. Po prenose informácií do všetkých oblastí mozgu, ktoré sú zodpovedné za naše vnímanie, nastáva skutočná zmyslová skúsenosť. Nociceptívna bolesť je relatívne jednoduchá reakcia na obzvlášť jednoduchý, akútny podnet. Ale mechanika zodpovedná za nociceptívnu bolesť nedokáže identifikovať javy, ako je bolesť, ktorá pretrváva napriek odstráneniu alebo vyliečeniu stimulácie, ako je napríklad fantómová bolesť končatín.
Bolesť a zápalová reakcia
Za okolností vážnejšieho poranenia, ako sú chirurgické rany, môže poškodenie tkaniva stimulovať zápalovú reakciu. Avšak iné stavy, najmä artritída, môžu byť tiež charakterizované pokračujúcimi prípadmi zápalu spojeného so symptómami intenzívnej bolesti. Mechanizmy tohto typu bolesti súvisiace s poškodením tkaniva a zápalovou odpoveďou sa líšia od nociceptívnej bolesti s včasným varovaním.
Pozorovaním rezu alebo miesta iného poškodenia alebo poranenia vzniká v nervovom systéme kaskáda hyperexcitabilných udalostí. Tento telesný fenomén „navíjania“ začína na koži, kde sa zosilňuje pozdĺž periférnych nervov a kulminuje pri reakcii z precitlivenosti pozdĺž miechy (dorzálny roh) a mozgu. Zápalové bunky potom obklopujú oblasti poškodenia tkaniva a tiež produkujú cytokíny a chemokíny, látky, ktoré majú sprostredkovať proces hojenia a regenerácie tkaniva. Ale tieto činidlá môžu byť tiež považované za dráždivé a upravujú vlastnosti primárnych senzorických neurónov obklopujúcich oblasť traumy.
Hlavné faktory, ktoré spúšťajú zápalovú bolesť, teda zahŕňajú poškodenie vysokoprahových nociceptorov, známe ako periférna senzibilizácia, zmeny a alterácie neurónov v nervovom systéme a zosilnenie excitability neurónov v CNS. To predstavuje centrálnu senzibilizáciu a je zodpovedné za hypersenzitivitu, kde oblasti susediace s oblasťami skutočného poranenia budú pociťovať bolesť, ako keby boli zranené. Tieto tkanivá môžu tiež reagovať na stimuláciu, ktorá za normálnych okolností nespôsobuje bolesť, ako je dotyk, nosenie oblečenia, mierny tlak alebo dokonca kefovanie vlastných vlasov, ako keby boli skutočne bolestivé, nazývané alodýnia.
Neuropatická bolesť je výsledkom poškodenia alebo poranenia nervového systému, ako je syndróm karpálneho tunela, postherpetická neuralgia a diabetická neuropatia. Hoci sa niektoré z mechanizmov, ktoré zrejme spôsobujú neuropatickú bolesť, prekrývajú s mechanizmami zodpovednými za zápalovú bolesť, mnohé z nich sú odlišné, a preto si budú vyžadovať odlišný prístup k ich liečbe.
Proces periférnej a centrálnej senzibilizácie je udržiavaný, aspoň teoreticky a experimentálne, počas excitačného neurotransmitera, glutamátu, o ktorom sa predpokladá, že sa uvoľňuje, keď je aktivovaný N-metyl-D-aspartátový (NMDA) receptor.
Nervový systém je tvorený buď inhibičnými alebo excitačnými neurotransmitermi. Väčšina z toho, čo nášmu nervovému systému umožňuje primerane reagovať na poškodenie alebo zranenie, je jemné ladenie alebo inhibícia rôznych procesov. Nadmerná excitácia nervového systému sa považuje za problém mnohých rôznych porúch. Napríklad nadmerná aktivácia NMDA receptora môže súvisieť aj s afektívnymi poruchami, sympatickými abnormalitami a dokonca toleranciou opiátov.
Dokonca aj obyčajná nociceptívna bolesť do určitej miery aktivuje NMDA receptor a predpokladá sa, že vedie k uvoľňovaniu glutamátu. Pri neuropatickej bolesti je však kľúčová precitlivenosť na NMDA receptor.
Pri iných typoch chronickej bolesti, ako je fibromyalgia a bolesti hlavy tenzného typu, môžu niektoré mechanizmy aktívne pri zápalovej a neuropatickej bolesti vytvárať podobné abnormality v systéme bolesti, vrátane centrálnej senzibilizácie, vyššej excitability somatosenzorických dráh a zníženia inhibičné mechanizmy centrálneho nervového systému.
Periférna senzibilizácia
Cyklooxygenáza (COX) tiež hrá zásadnú funkciu pri periférnej aj centrálnej senzibilizácii. COX-2 je jedným z enzýmov, ktoré sú indukované počas zápalového procesu; COX-2 premieňa kyselinu arachidónovú na prostaglandíny, ktoré zvyšujú citlivosť periférnych nociceptorových zakončení. V podstate periférny zápal tiež spôsobuje produkciu COX-2 z CNS. Signály z periférnych nociceptorov sú čiastočne zodpovedné za túto upreguláciu, ale zdá sa, že existuje aj humorálna zložka prenosu signálov bolesti cez hematoencefalickú bariéru.
Napríklad v experimentálnych modeloch sa COX-2 generuje z CNS, aj keď zvieratá dostanú blokádu senzorických nervov pred periférnou zápalovou stimuláciou. COX-2, ktorý je exprimovaný nad neurónmi dorzálneho rohu miechy, uvoľňuje prostaglandíny, ktoré pôsobia na centrálne zakončenia alebo presynaptické zakončenia nociceptívnych senzorických vlákien, aby sa zvýšilo uvoľňovanie vysielača. Okrem toho pôsobia postsynapticky na neuróny dorzálneho rohu, čím vytvárajú priamu depolarizáciu. A nakoniec, inhibujú aktivitu glycínového receptora, a to je inhibičný transmiter. Preto prostaglandíny spôsobujú zvýšenie excitability centrálnych neurónov.
Plasticita mozgu a centrálna senzibilizácia
Centrálna senzibilizácia popisuje zmeny, ku ktorým dochádza v mozgu ako reakcia na opakovanú nervovú stimuláciu. Po opakovaných stimuloch sa množstvo hormónov a mozgových elektrických signálov mení, pretože neuróny si vytvárajú „pamäť“ na reakciu na tieto príznaky. Neustála stimulácia vytvára výkonnejšiu mozgovú pamäť, takže mozog bude reagovať rýchlejšie a efektívnejšie, keď podstúpi rovnakú stimuláciu v budúcnosti. Následné modifikácie v mozgovom zapojení a reakcii sa označujú ako nervová plasticita, ktorá opisuje schopnosť mozgu ľahko sa meniť alebo centrálna senzibilizácia. Preto je mozog aktivovaný alebo senzibilizovaný predchádzajúcimi alebo opakovanými stimulmi, aby sa stal vzrušujúcejším.
Kolísanie centrálnej senzibilizácie nastáva po opakovaných stretnutiach s bolesťou. Výskum na zvieratách naznačuje, že opakované vystavenie bolestivej stimulácii zmení prah bolesti zvieraťa a vedie k silnejšej reakcii na bolesť. Vedci si myslia, že tieto úpravy môžu vysvetliť pretrvávajúcu bolesť, ktorá sa môže vyskytnúť aj po úspešnej operácii chrbta. Hoci herniovaný disk môže byť odstránený zo zovretého nervu, bolesť môže pokračovať ako spomienka na kompresiu nervu. Novorodenci podstupujúci obriezku bez anestézie budú hlbšie reagovať na budúcu bolestivú stimuláciu, ako sú bežné injekcie, očkovania a iné bolestivé procesy. Tieto deti majú nielen vyššiu hemodynamickú reakciu, známu ako tachykardia a tachypnoe, ale rozvinie sa u nich aj zvýšený plač.
Táto neurologická pamäť bolesti bola dôkladne študovaná. V správe o svojich predchádzajúcich výskumných štúdiách Woolf poznamenal, že zlepšená reflexná excitabilita po poškodení alebo poranení periférneho tkaniva nezávisí od pokračujúcich periférnych vstupných signálov; skôr hodiny po periférnej traume sa receptorové polia neurónov chrbtového rohu chrbtice naďalej zväčšovali. Výskumníci tiež zdokumentovali význam spinálneho NMDA receptora pre vyvolanie a udržiavanie centrálnej senzibilizácie.
Význam pre liečbu bolesti
Akonáhle sa vytvorí centrálna senzibilizácia, na jej potlačenie sú často potrebné väčšie dávky analgetík. Preventívna analgézia alebo terapia pred progresiou bolesti môže znížiť účinky všetkých týchto stimulácií na CNS. Woolfová preukázala, že dávka morfínu potrebná na zastavenie centrálnej hyperexcitability, podaná pred krátkou škodlivou elektrickou stimuláciou u potkanov, bola jedna desatina dávky potrebnej na zrušenie aktivity po jej vyrastení. To sa premieta do klinickej praxe.
V klinickej štúdii so 60 pacientkami, ktoré podstúpili abdominálnu hysterektómiu, jedinci, ktorí dostali 10 mg morfínu intravenózne v čase úvodu do anestézie, potrebovali na kontrolu pooperačnej bolesti výrazne menej morfínu. Okrem toho citlivosť na bolesť okolo rany, označovaná ako sekundárna hyperalgézia, bola tiež znížená v skupine vopred ošetrenej morfínom. Preemptívna analgézia sa použila s porovnateľným úspechom v rade chirurgických nastavení, vrátane predspinálnej operácie a postortopedickej operácie.
Jednorazová dávka 40 alebo 60 mg/kg rektálneho acetaminofénu má jasný morfín šetriaci účinok pri jednodňovej chirurgii u detí, ak sa podáva na úvod do anestézie. Okrem toho deti s dostatočnou analgéziou s acetaminofénom zaznamenali výrazne menej pooperačnej nevoľnosti a vracania.
Antagonisty NMDA receptora poskytujú pooperačnú analgéziu, keď sa podávajú predoperačne. V literatúre existujú rôzne správy podporujúce použitie ketamínu a dextrometorfánu v predoperačnom období. U pacientov, ktorí podstúpili rekonštrukciu predného skríženého väzu, bola 24-hodinová pacientom kontrolovaná spotreba opiátov z analgézie významne nižšia v predoperačnej kategórii dextrometorfánu oproti skupine s placebom.
V dvojito zaslepených, placebom kontrolovaných výskumných štúdiách bol gabapentín indikovaný ako premedikačné analgetikum u pacientov podstupujúcich mastektómiu a hysterektómiu. Predoperačný perorálny gabapentín znížil skóre bolesti a pooperačnú spotrebu analgetík bez medzery vo vedľajších účinkoch v porovnaní s placebom.
Predoperačné podávanie nesteroidných protizápalových liekov (NSAID) preukázalo významný pokles užívania opiátov po operácii. COX-2 sú vhodnejšie kvôli ich relatívnemu nedostatku účinkov na krvné doštičky a významnému profilu gastrointestinálnej bezpečnosti v porovnaní s konvenčnými NSAID. Celecoxib, rofekoxib, valdekoxib a parekoxib podávané pred operáciou mimo USA znižujú pooperačné užívanie narkotík o viac ako 40 percent, pričom mnohí pacienti užívajú menej ako polovicu opioidov v porovnaní s placebom.
Zdá sa, že blokovanie nervového vedenia v predoperačnom období zabraňuje rozvoju centrálnej senzibilizácie. Syndróm fantómových končatín (PLS) sa pripisuje fenoménu navíjania chrbtice.�Pacienti s amputáciou
často majú odstránenú pálivú alebo mravenčiu bolesť v časti tela. Jednou z možných príčin je, že nervové vlákna na pahýľi sú stimulované a mozog interpretuje signály ako pochádzajúce z amputovanej časti. Druhým je preskupenie v kortikálnych oblastiach tak, že oblasť povedzme pre ruku teraz reaguje na signály z iných častí tela, ale stále ich interpretuje ako prichádzajúce pre amputovanú ruku.
Avšak u pacientov, ktorí podstúpili amputáciu dolných končatín v epidurálnej anestézii, sa u jedného z 11 pacientov, ktorí dostávali lumbálnu epidurálnu blokádu bupivakaínom a morfínom počas 72 hodín pred operáciou, nevyvinula PLS. U ľudí, ktorí podstúpili celkovú anestéziu bez predchádzajúcej lumbálnej epidurálnej blokády, malo 5 zo 14 pacientov PLS po 6 týždňoch au 3 pokračovali v PLS po 1 roku.
Woolf a Chong poznamenali, že dokonalá predoperačná, intraoperačná a pooperačná liečba zahŕňa „NSAID na zníženie aktivácie/centralizácie nociceptorov, lokálne anestetiká na blokovanie senzorického prítoku a centrálne pôsobiace lieky, ako sú opiáty“. Zníženie perioperačnej bolesti pomocou preventívnych techník zvyšuje spokojnosť, urýchľuje výtok, šetrí užívanie opioidov spolu so zníženou zápchou, sedáciou, nevoľnosťou a retenciou moču a môže dokonca zastaviť rozvoj chronickej bolesti. Anestéziológovia a chirurgovia by mali zvážiť integráciu týchto techník do svojej každodennej praxe.
Keď sa bolesť objaví v dôsledku poškodenia alebo poranenia v dôsledku chirurgického zákroku, miecha môže dosiahnuť hyperexcitabilný stav, v ktorom dochádza k nadmerným bolestivým reakciám, ktoré môžu pretrvávať niekoľko dní, týždňov alebo dokonca rokov.
Prečo má lokalizované zranenie v dôsledku traumy u niektorých pacientov chronickú, nezvládnuteľnú bolesť? Poškodenie tkaniva vedie ku konštelácii zmien v dráždivosti chrbtice, vrátane zvýšeného spontánneho výboja, väčšej amplitúdy a dĺžky odozvy, zníženého prahu, zvýšeného výboja pri opakovanej stimulácii a rozšírených receptívnych polí. Pretrvávanie týchto zmien, ktoré sa súhrnne nazývajú centrálna senzibilizácia, sa javí ako zásadné pre predĺžené zvýšenie citlivosti na bolesť, ktorá definuje chronickú bolesť. Početné lieky a/alebo lieky, ako aj lokálna anestetická neurálna blokáda môžu obmedziť veľkosť vetrania centrálneho nervového systému (CNS), čo dokazuje znížená bolesť a znížená spotreba opioidov v preventívnych analgetických modeloch.
Pohľad Dr. Alexa Jimeneza
Chiropraktická starostlivosť je alternatívna možnosť liečby, ktorá využíva úpravy chrbtice a ručné manipulácie na bezpečné a efektívne obnovenie, ako aj na udržanie správneho zarovnania chrbtice. Výskumné štúdie zistili, že nesprávne postavenie chrbtice alebo subluxácia môže viesť k chronickej bolesti. Chiropraktická starostlivosť sa bežne využíva na zvládanie bolesti, aj keď symptómy nie sú spojené s poranením a/alebo stavom v muskuloskeletálnom a nervovom systéme. Opatrným opätovným vyrovnaním chrbtice sa a chiropraktik môže pomôcť znížiť stres a tlak zo štruktúr obklopujúcich hlavnú zložku základov tela, čo v konečnom dôsledku poskytuje úľavu od bolesti.
Funkcia a bolesť enterického nervového systému
Pokiaľ ide o obmedzené používanie liekov a/alebo liekov, vrátane opioidov, aby sa predišlo vedľajším účinkom, ako sú gastrointestinálne zdravotné problémy, môže hrať úlohu správna funkcia enterálneho nervového systému.
Enterický nervový systém (ENS) alebo vnútorný nervový systém je jednou z kľúčových vetiev autonómneho nervového systému (ANS) a pozostáva zo sieťovitého systému nervov, ktorý moduluje úlohu gastrointestinálneho traktu. Je schopný pôsobiť nezávisle od sympatického a parasympatického nervového systému, aj keď môže byť nimi ovplyvnený. ENS možno nazvať aj druhým mozgom.�Pochádza z buniek neurálnej lišty.
Enterický nervový systém u ľudí sa skladá z približne 500 miliónov neurónov, vrátane mnohých typov Dogielových buniek, čo je približne jedna dvestotina množstva neurónov v mozgu. Enterický nervový systém je vložený do výstelky gastrointestinálneho systému, počínajúc pažerákom a siahajúcim až po konečník. Dogielove bunky, tiež známe ako Dogielove bunky, označujú nejaký druh multipolárnych nadobličkových tkanív v prevertebrálnych sympatických gangliách.
ENS je schopná vykonávať autonómne funkcie, ako je koordinácia reflexov; Aj keď dostáva značnú inerváciu v autonómnom nervovom systéme, funguje a môže fungovať nezávisle od mozgu a miechy.�Črevný nervový systém bol opísaný ako „druhý mozog“ z viacerých dôvodov. Enterický nervový systém môže fungovať autonómne. Normálne komunikuje s centrálnym nervovým systémom (CNS) cez parasympatikus alebo cez nervus vagus a so sympatikom, to znamená cez prevertebrálne gangliá, nervový systém. Štúdie na stavovcoch však odhaľujú, že keď je blúdivý nerv prerušený, enterický nervový systém naďalej funguje.
U stavovcov enterický nervový systém zahŕňa eferentné neuróny, aferentné neuróny a interneuróny, z ktorých všetky robia črevný nervový systém schopným niesť reflexy a pôsobiť ako integrujúce centrum v neprítomnosti vstupu CNS. Senzorické neuróny hlásia mechanické a chemické podmienky. Enterický nervový systém má schopnosť meniť svoju odpoveď na základe takých faktorov, ako je zloženie živín a objemové zloženie. Okrem toho ENS obsahuje podporné bunky, ktoré sú veľmi podobné astroglii mozgu a difúznu bariéru okolo kapilár obklopujúcich gangliá, ktorá je ako hematoencefalická bariéra krvných ciev.
Enterický nervový systém (ENS) hrá kľúčovú úlohu v zápalových a nociceptívnych procesoch. Lieky a/alebo lieky, ktoré interagujú s ENS nedávno vzbudili značný záujem kvôli ich schopnosti regulovať mnohé aspekty fyziológie a patofyziológie čreva. Najmä experimenty na zvieratách ukázali, že receptory aktivované proteinázou (PAR) môžu byť nevyhnutné pre neurogénny zápal v čreve. Okrem toho sa zdá, že agonisty PAR2 vyvolávajú črevnú hypersenzitivitu a hyperalgetické stavy, čo naznačuje úlohu tohto receptora pri vnímaní viscerálnej bolesti.
Okrem toho PAR spolu s proteinázami, ktoré ich aktivujú, predstavujú vzrušujúce nové ciele pre terapeutickú intervenciu na ENS. Rozsah našich informácií je obmedzený na chiropraktiku, ako aj na poranenia a stavy chrbtice. Ak chcete prediskutovať túto tému, neváhajte sa opýtať Dr. Jimeneza alebo nás kontaktujte na adrese 915-850-0900 .
Kurátorom je Dr. Alex Jimenez
Ďalšie témy: Ischias
ischias sa v medicíne označuje ako súbor symptómov, a nie ako jediné zranenie a/alebo stav. Symptómy bolesti sedacieho nervu alebo ischias sa môžu líšiť frekvenciou a intenzitou, najčastejšie sa však opisuje ako náhla, ostrá (nožovitá) alebo elektrická bolesť, ktorá vyžaruje z dolnej časti chrbta do zadku, bokov, stehien a nohy do chodidla. Ďalšie príznaky ischias môžu zahŕňať pocity brnenia alebo pálenia, necitlivosť a slabosť po celej dĺžke sedacieho nervu. Ischias najčastejšie postihuje jedincov vo veku 30 až 50 rokov. Často sa môže vyvinúť ako dôsledok degenerácie chrbtice vekom, avšak stlačenie a podráždenie sedacieho nervu spôsobené vydutím resp. herniovaný disk, okrem iných zdravotných problémov chrbtice, môže tiež spôsobiť bolesť sedacieho nervu.
Nástroj Find A Practitioner od IFM je najväčšou sieťou odporúčaní vo funkčnej medicíne, ktorá bola vytvorená s cieľom pomôcť pacientom nájsť praktických lekárov kdekoľvek na svete. IFM Certified Practitioners sú vo výsledkoch vyhľadávania uvedení na prvom mieste vzhľadom na ich rozsiahle vzdelanie v oblasti funkčnej medicíny

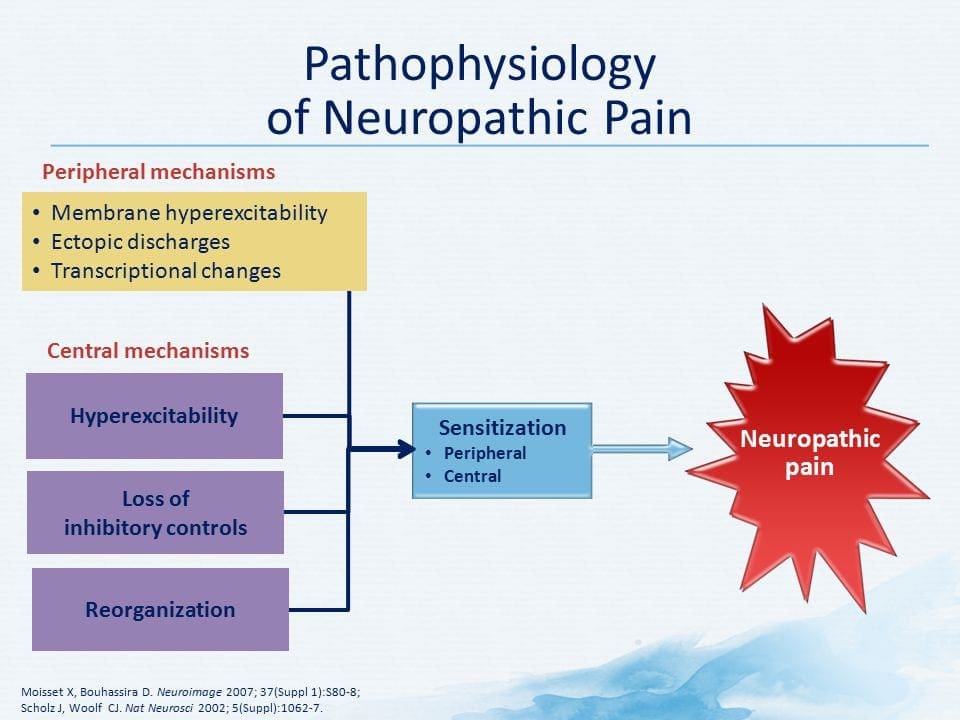
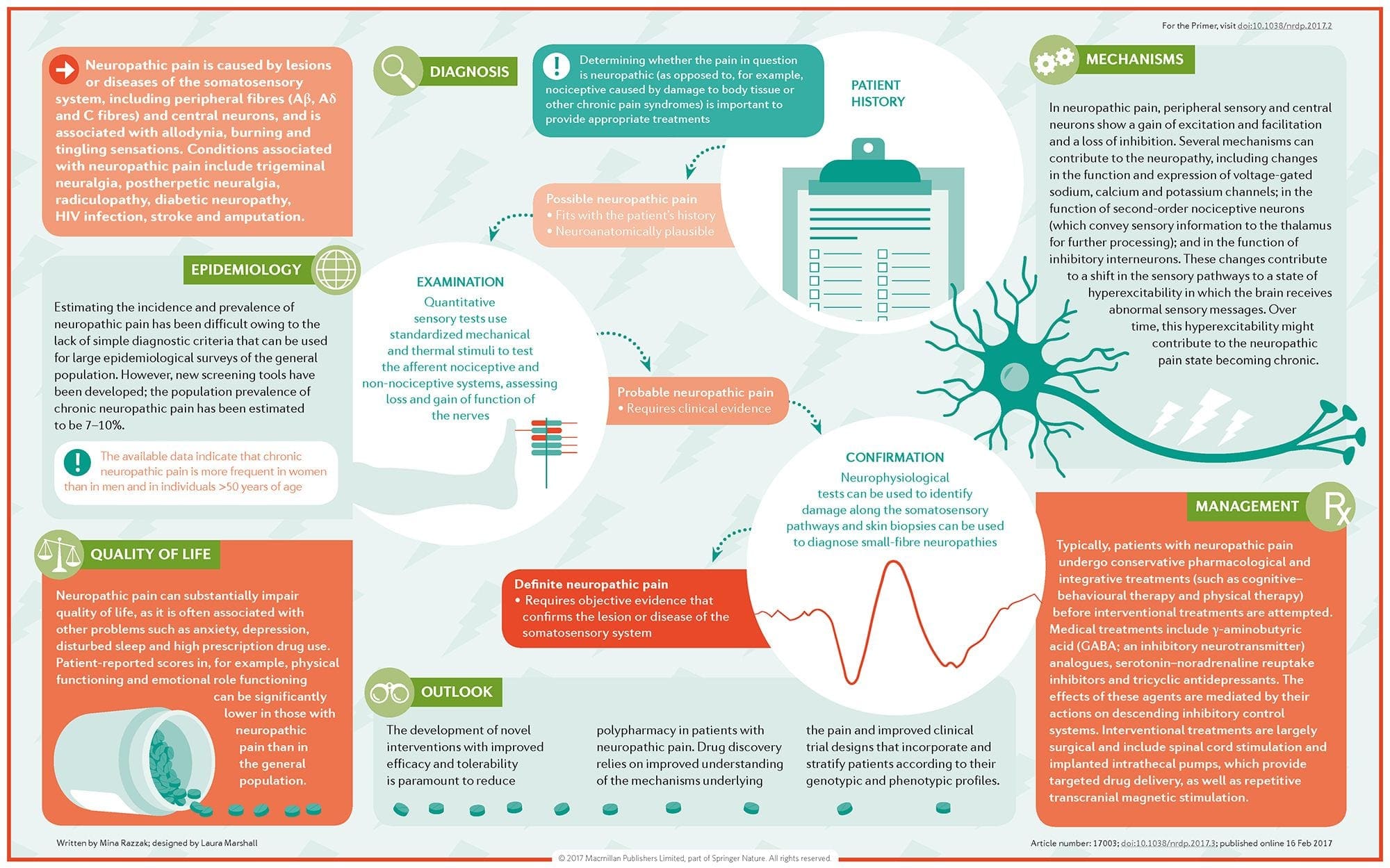




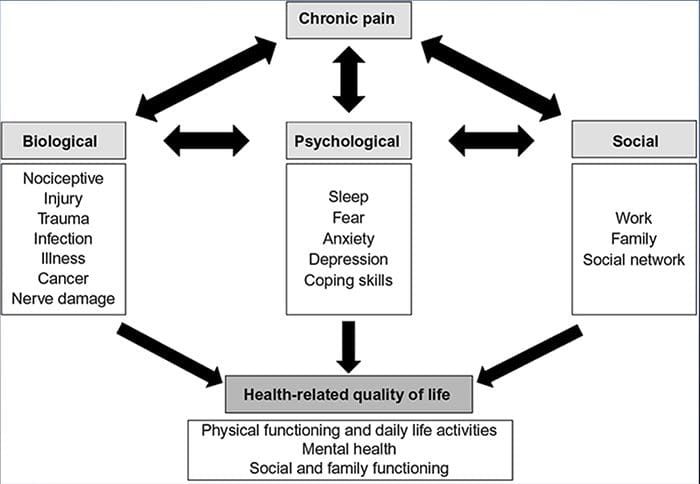
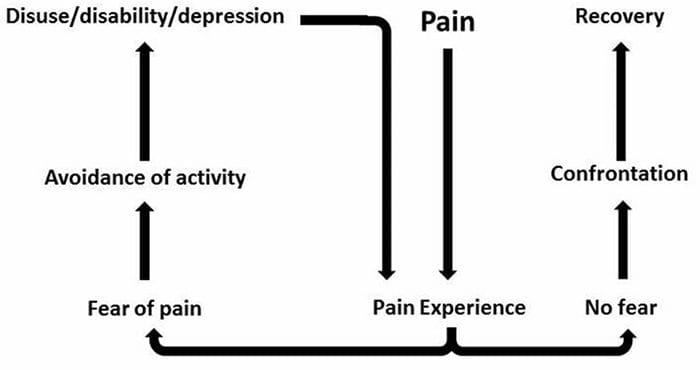
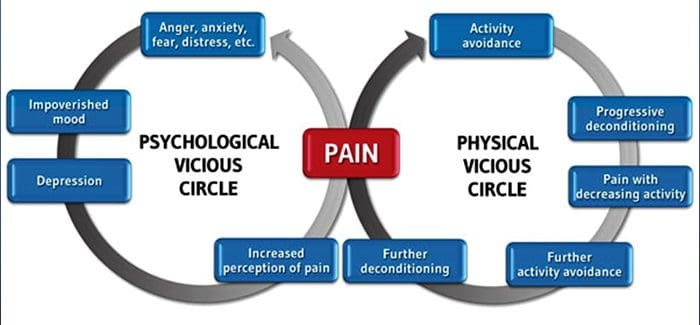
















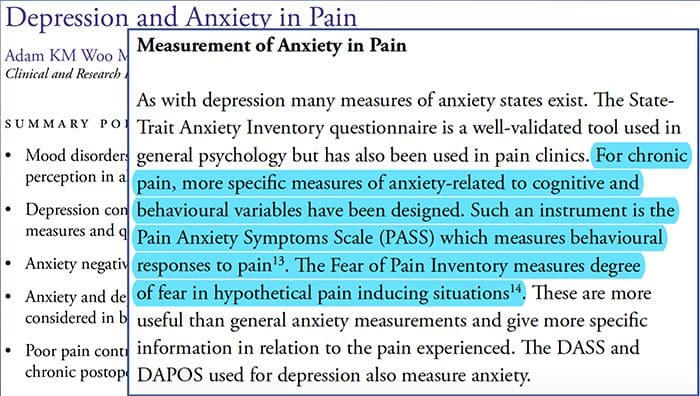


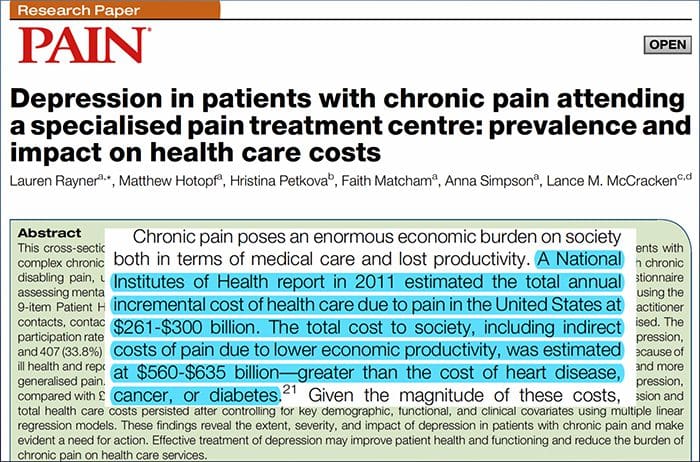



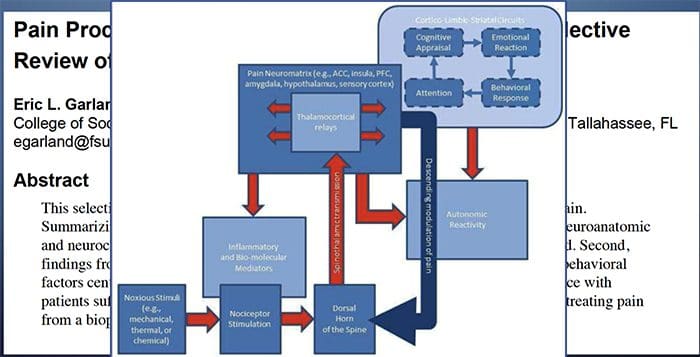






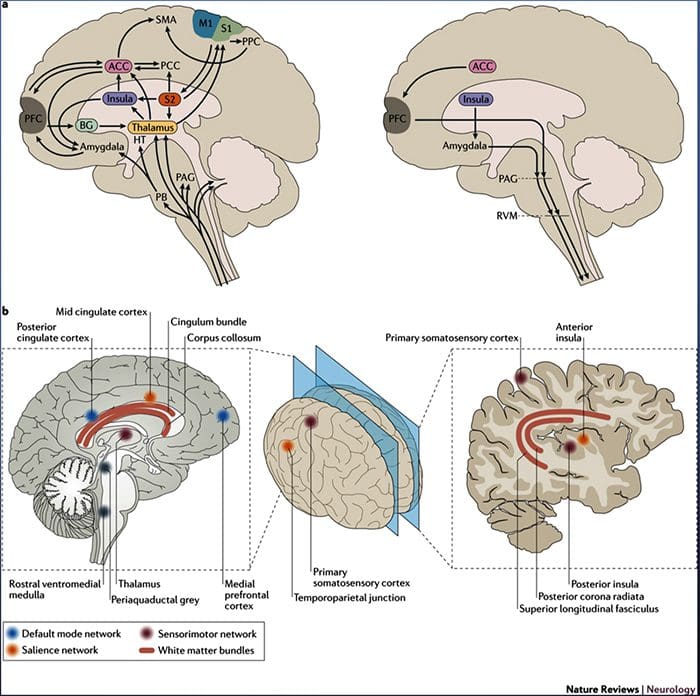

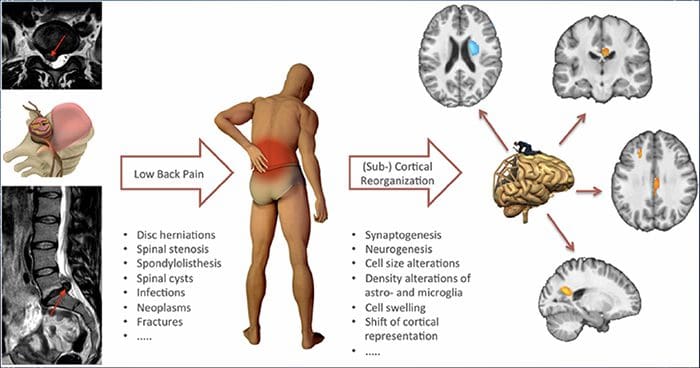

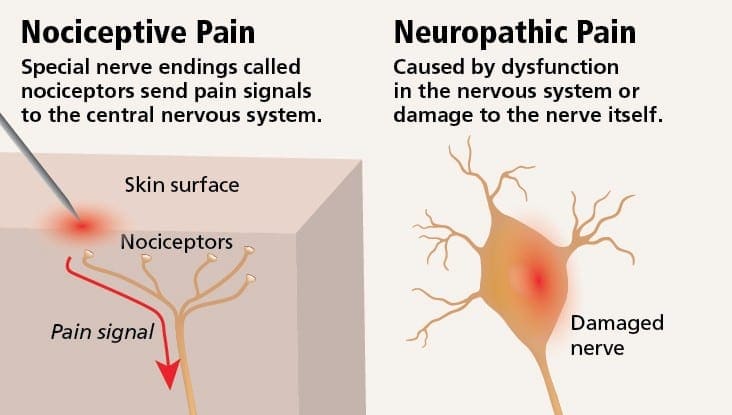
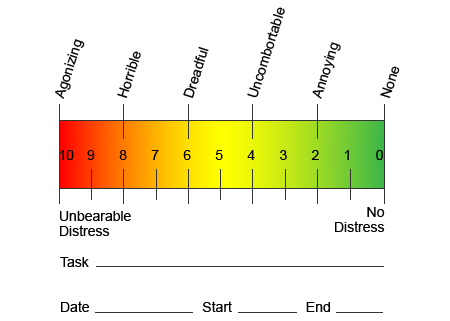


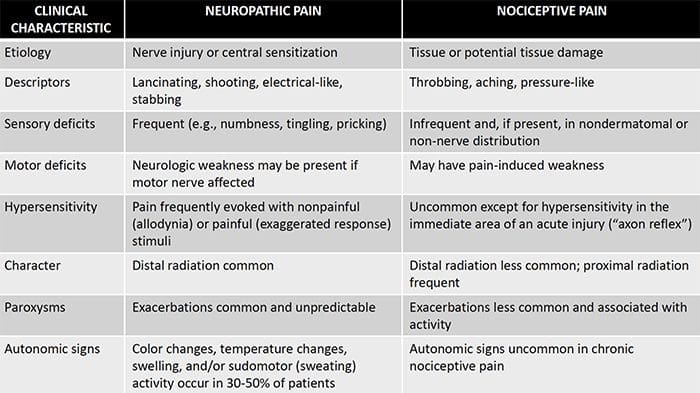
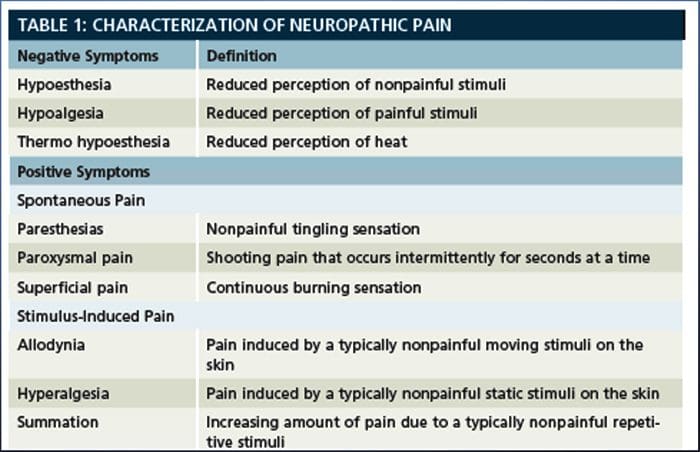
 PATOGENÉZA NEUROPATICKEJ BOLESTI
PATOGENÉZA NEUROPATICKEJ BOLESTI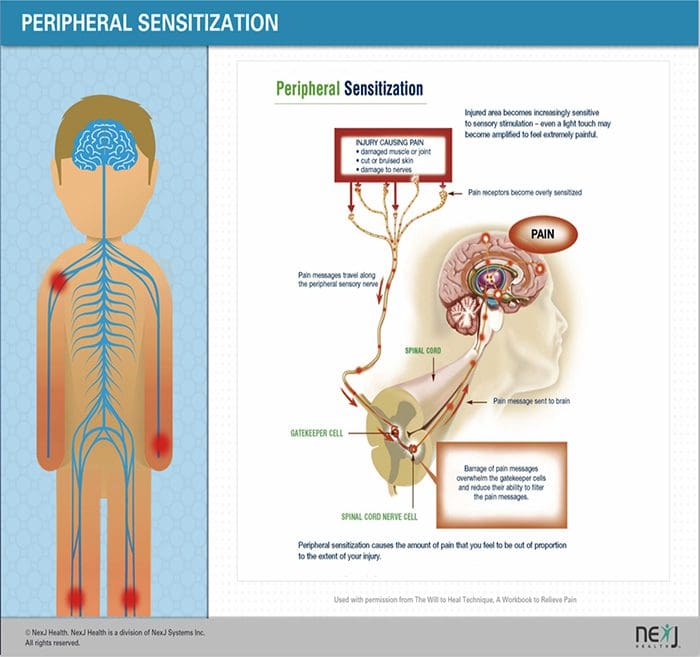
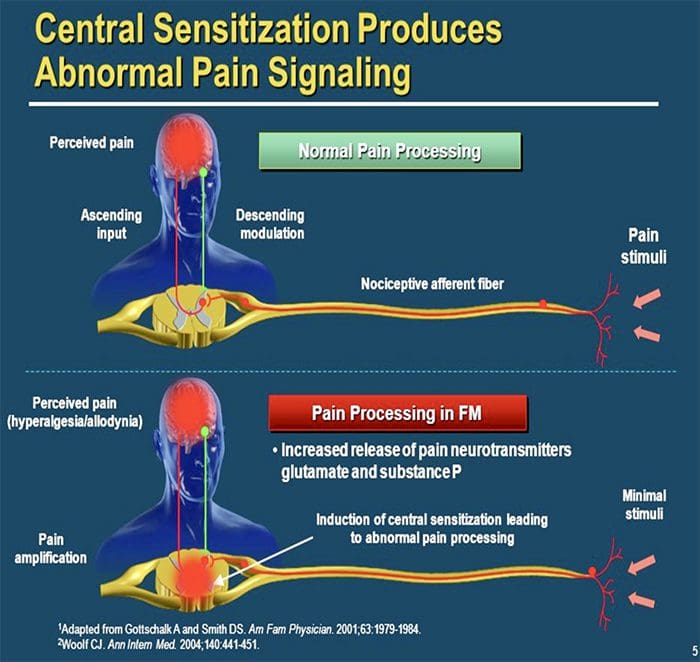
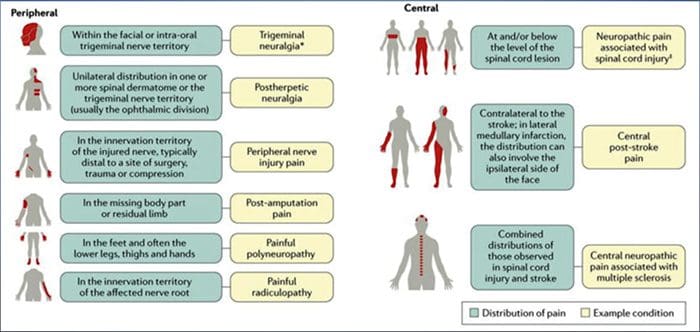 BEŽNÉ PRÍČINY
BEŽNÉ PRÍČINY
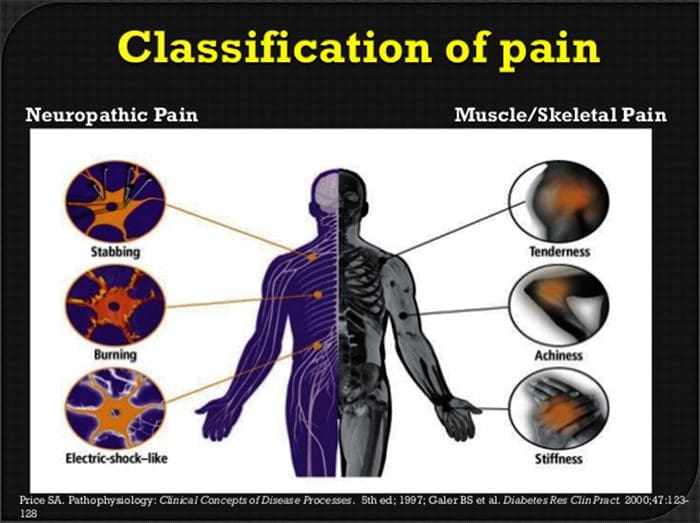
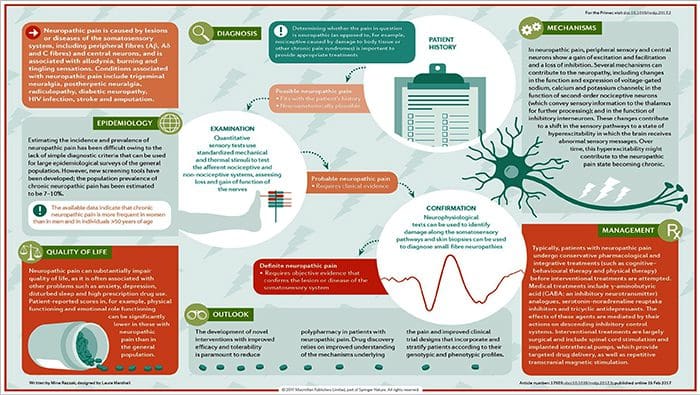
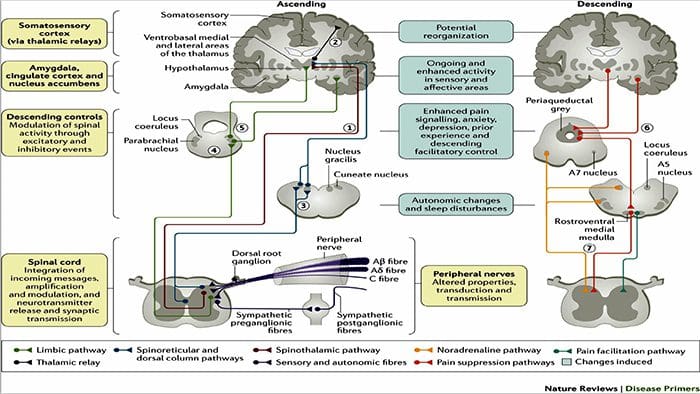
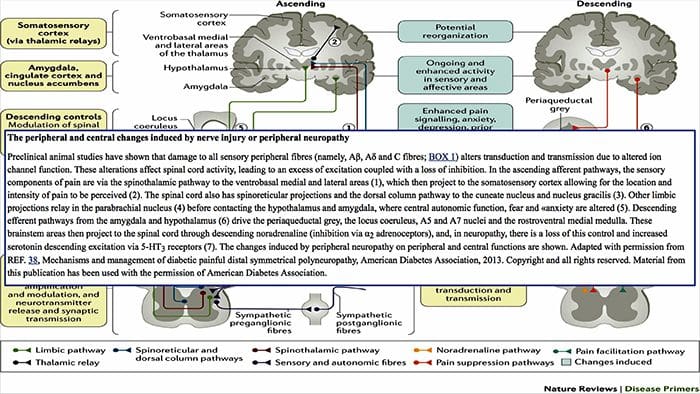

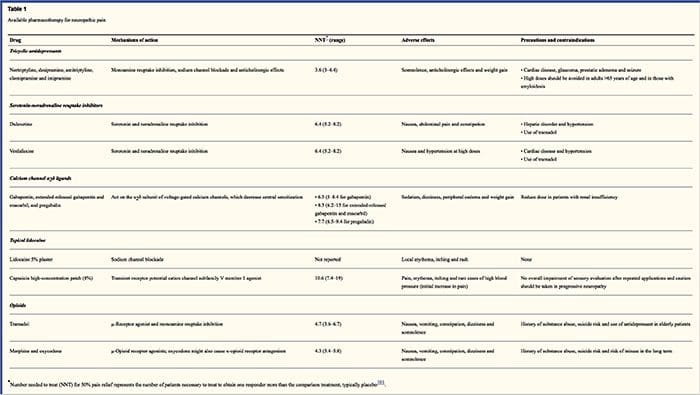




 FANTÓMOVÁ BOLESŤ KONČATINY A ROZŠÍRENÁ REALITA
FANTÓMOVÁ BOLESŤ KONČATINY A ROZŠÍRENÁ REALITA
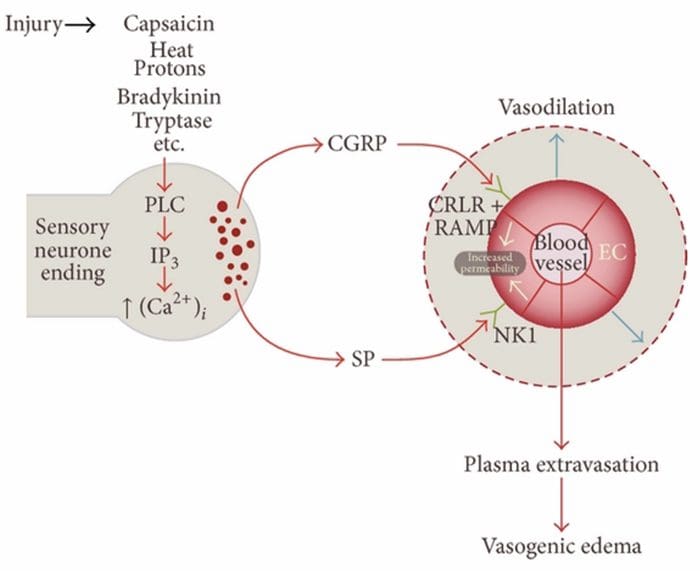 BEŽNÉ PRÍČINY
BEŽNÉ PRÍČINY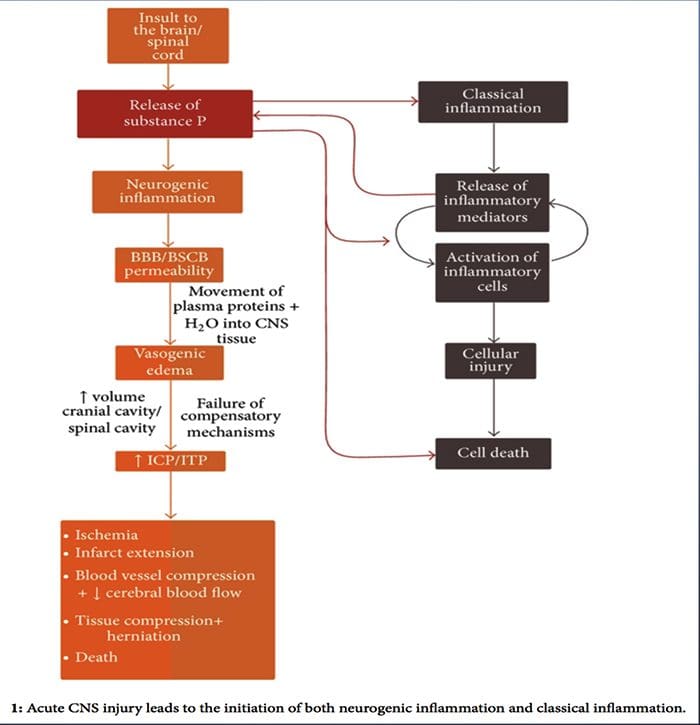


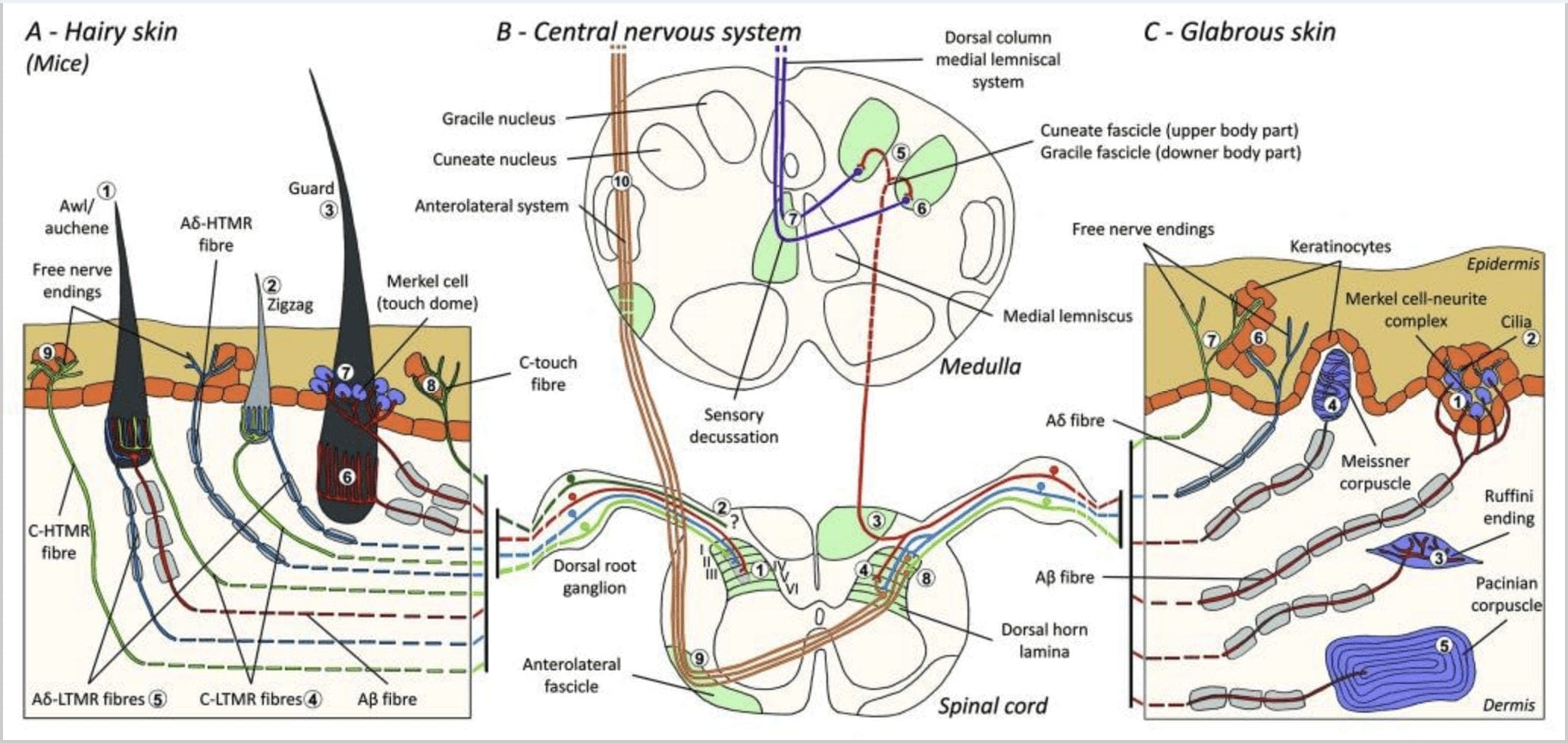
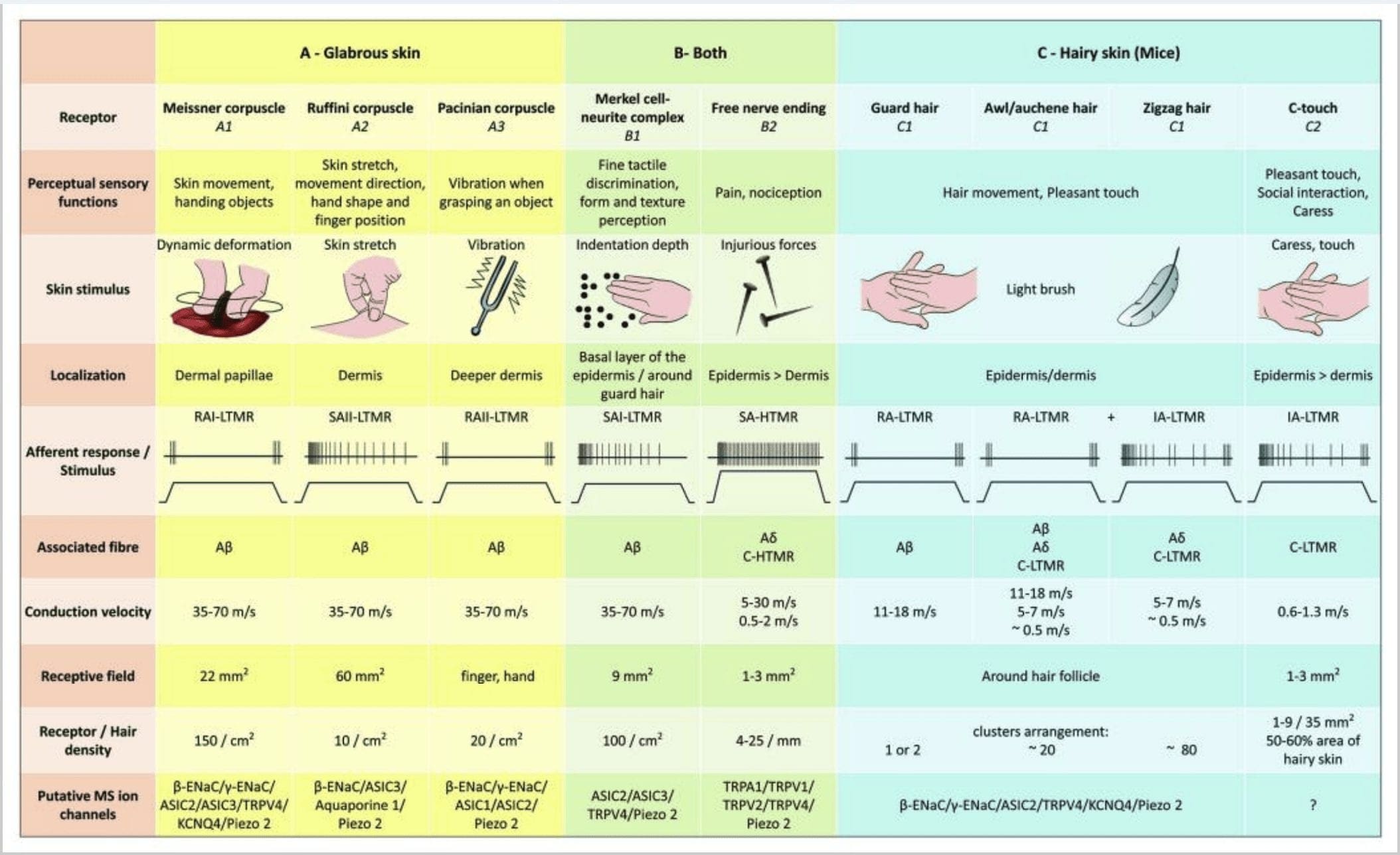

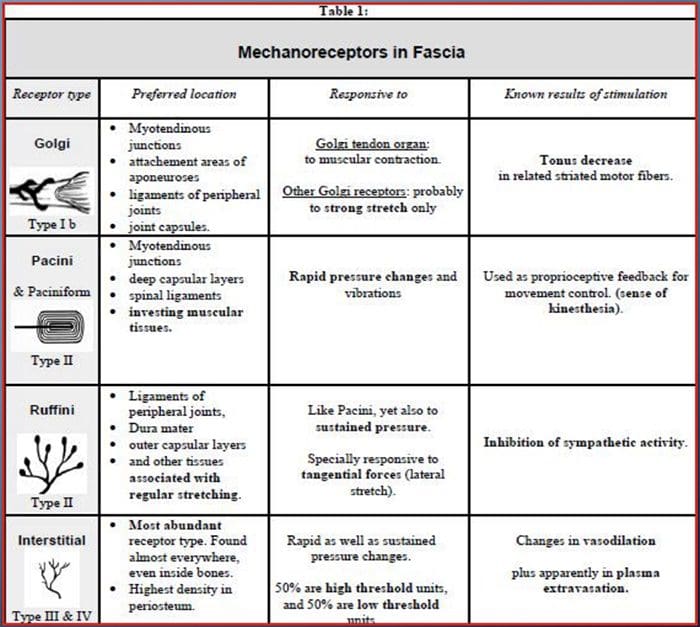 TEÓRIA RIADENIA BRÁNY BOLESTI
TEÓRIA RIADENIA BRÁNY BOLESTI
 CHRBTOVÝ STĹPEC MEDIÁLNA LEMNISKÁLNA DRÁHA
CHRBTOVÝ STĹPEC MEDIÁLNA LEMNISKÁLNA DRÁHA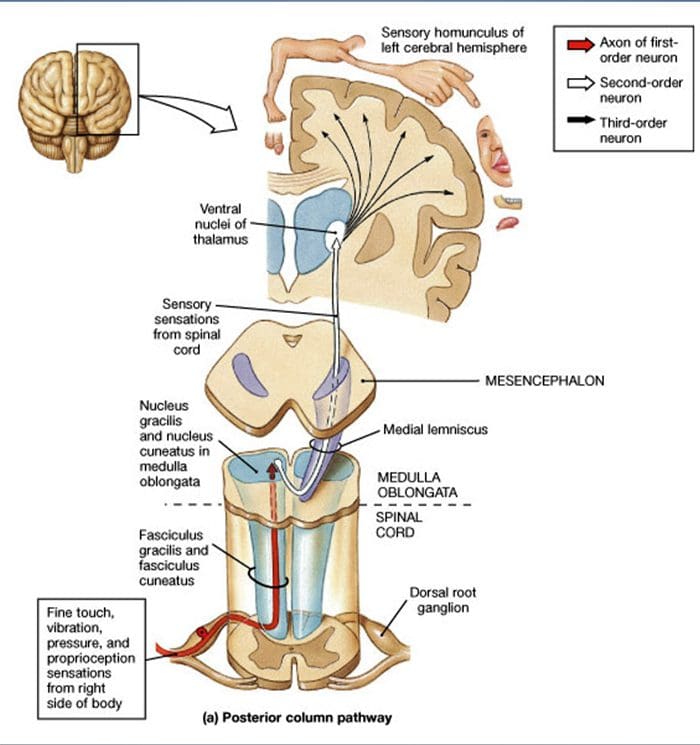 VYUŽITIE PERIFERNÝCH MECHANORECEPTOROV NA ZMENU VNÍMANIA BOLESTI
VYUŽITIE PERIFERNÝCH MECHANORECEPTOROV NA ZMENU VNÍMANIA BOLESTI
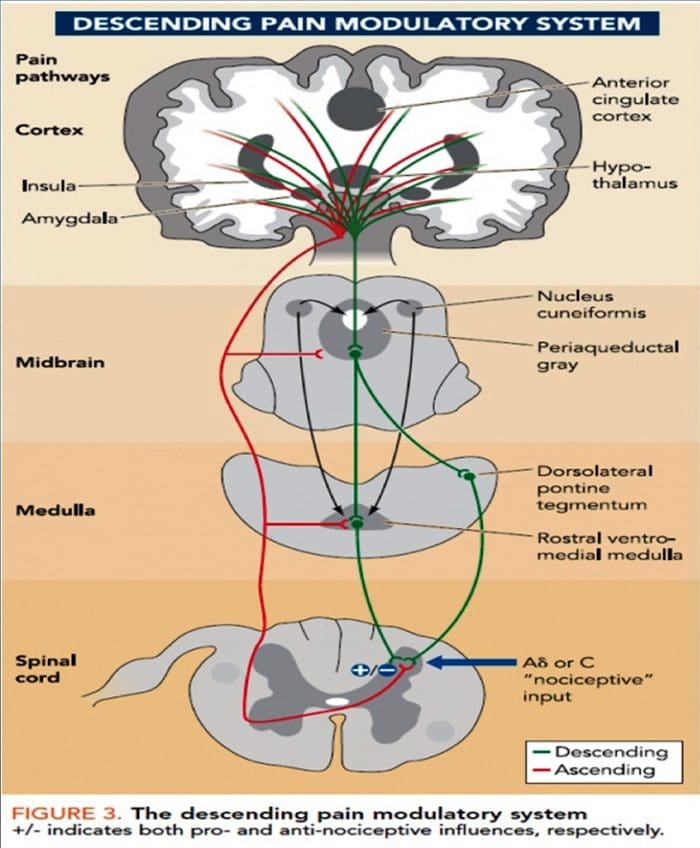
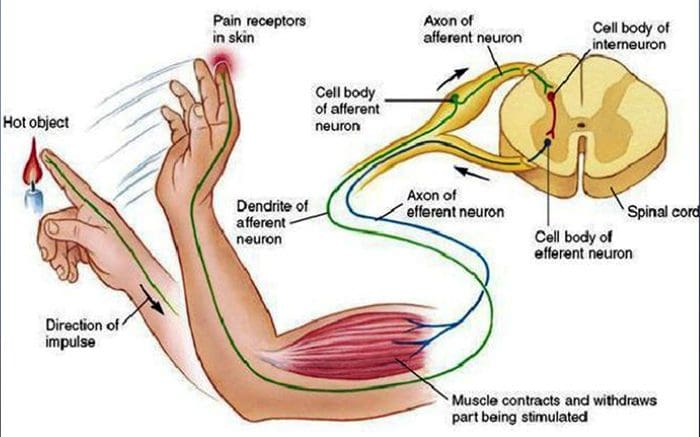
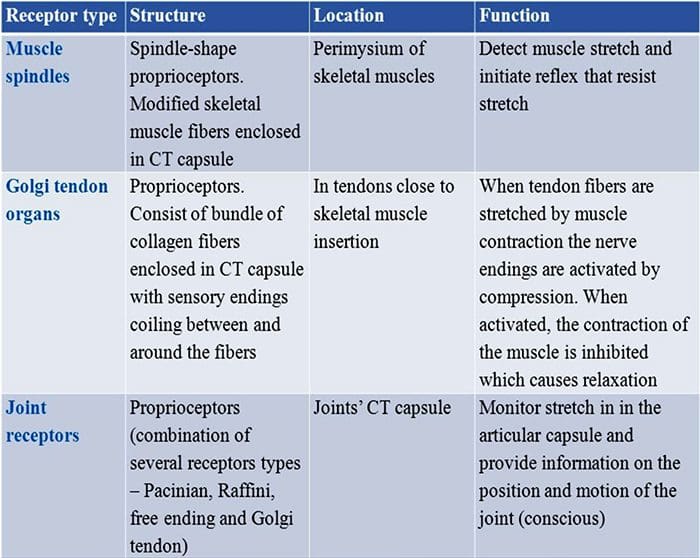
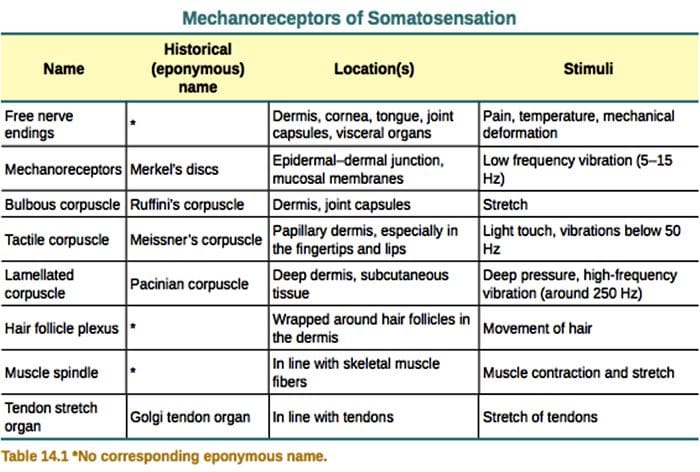 VIBRÁCIE
VIBRÁCIE
 ĽAHKÝ DOTYK
ĽAHKÝ DOTYK



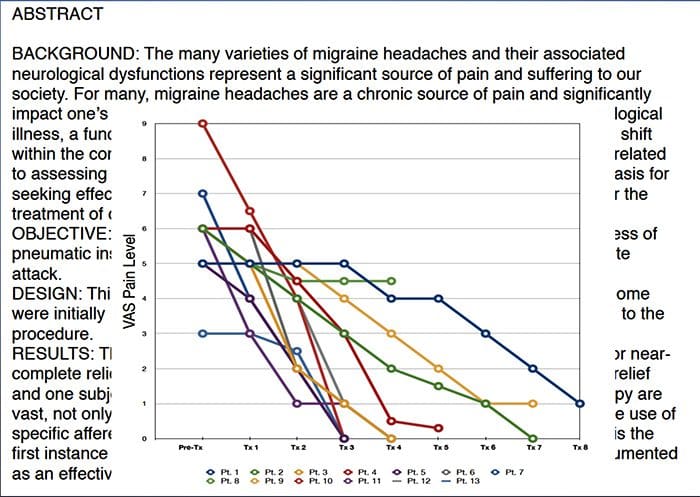


 CASE STUDY
CASE STUDY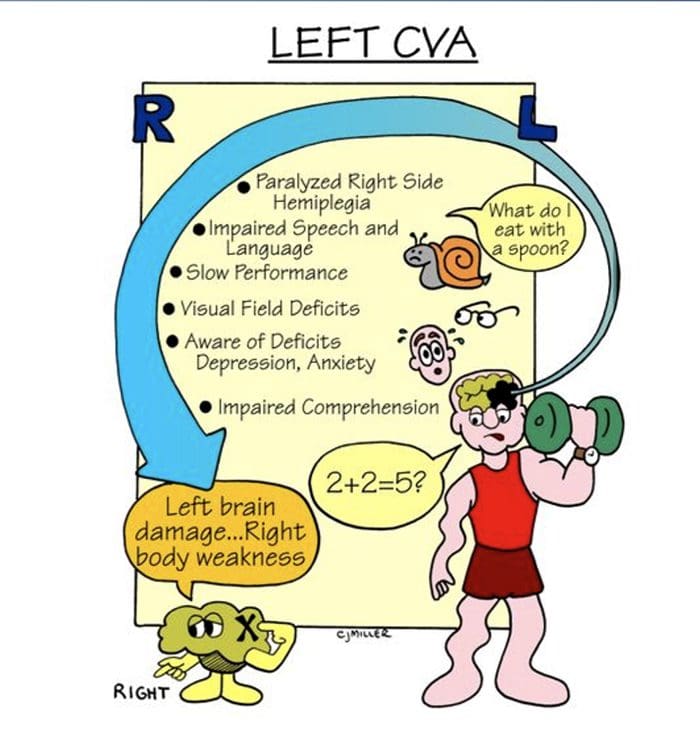 NAJLEPŠIE FYZICKÉ VYŠETRENIE
NAJLEPŠIE FYZICKÉ VYŠETRENIE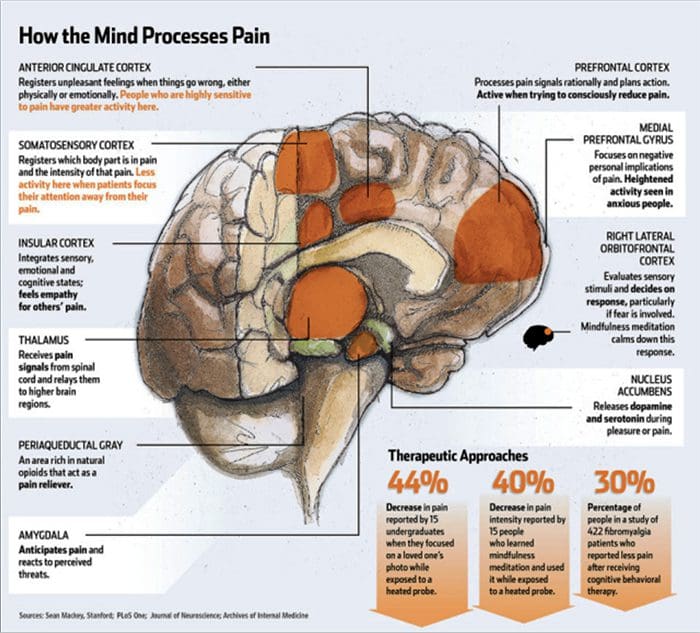 NAJLEPŠIE FYZICKÉ VYŠETRENIE
NAJLEPŠIE FYZICKÉ VYŠETRENIE
 PO DVOCH DŇOCH
PO DVOCH DŇOCH