Chiropraxe a tím funkčnej medicíny Back Clinic pre oxidačný stres. Oxidačný stres je definovaný ako narušenie rovnováhy medzi produkciou reaktívneho kyslíka (voľné radikály) a antioxidačnou obranou. Inými slovami, ide o nerovnováhu medzi produkciou voľných radikálov a schopnosťou tela pôsobiť proti škodlivým účinkom alebo ich detoxikovať prostredníctvom neutralizácie antioxidantmi. Oxidačný stres vedie k mnohým patofyziologickým stavom v tele. Patria sem neurodegeneratívne ochorenia, tj Parkinsonova choroba, Alzheimerova choroba, génové mutácie, rakovina, syndróm chronickej únavy, syndróm fragilného X, poruchy srdca a krvných ciev, ateroskleróza, srdcové zlyhanie, srdcový infarkt a zápalové ochorenia. K oxidácii dochádza za niekoľkých okolností:
bunky využívajú glukózu na výrobu energie
imunitný systém bojuje proti baktériám a vytvára zápal
telo detoxikuje škodliviny, pesticídy a cigaretový dym
V našom tele prebiehajú v každom okamihu milióny procesov, ktoré môžu viesť k oxidácii. Tu je niekoľko príznakov:
únava
Strata pamäti alebo mozgová hmla
Bolesť svalov a/alebo kĺbov
Vrásky spolu so sivými vlasmi
Znížený zrak
Bolesti hlavy a citlivosť na hluk
Citlivosť na infekcie
Výber organických potravín a vyhýbanie sa toxínom vo vašom prostredí je veľký rozdiel. To spolu so znížením stresu môže byť prospešné pri znižovaní oxidácie.
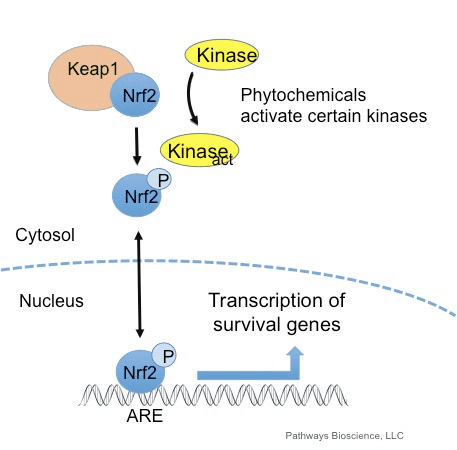
Oxidanty sa vo všeobecnosti vyrábajú kontrolovaným spôsobom, aby sa regulovali základné procesy v ľudskom tele, vrátane bunkového delenia, zápalu, imunitnej funkcie, autofágie a stresovej reakcie. K tomu však môže prispieť nekontrolovaná produkcia týchto oxidantov oxidačný stres, čo môže ovplyvniť bunkovú funkciu, čo vedie k rozvoju toxicity, chronických ochorení a rakoviny. Ochranné antioxidačné mechanizmy ľudského tela sú regulované sériou životne dôležitých dráh, ktoré riadia odpoveď bunky na oxidanty. Nukleárny faktor erytroidný 2-príbuzný faktor, inak známy ako Nrf2, je novým regulátorom bunkovej rezistencie voči oxidantom. Účelom nižšie uvedeného článku je diskutovať a demonštrovať vznikajúcu úlohu Nrf2 v mitochondriálnej funkcii.
abstraktné
Transkripčný faktor NF-E2 faktor 45 súvisiaci s p2 (Nrf2; názov génu NFE2L2) umožňuje adaptáciu a prežitie v podmienkach stresu reguláciou génovej expresie rôznych sietí cytoprotektívnych proteínov, vrátane antioxidačných, protizápalových a detoxikačných enzýmov. ako proteíny, ktoré pomáhajú pri oprave alebo odstraňovaní poškodených makromolekúl. Nrf2 má kľúčovú úlohu pri udržiavaní bunkovej redoxnej homeostázy reguláciou biosyntézy, využitia a regenerácie glutatiónu, tioredoxínu a NADPH a riadením produkcie reaktívnych foriem kyslíka mitochondriami a NADPH oxidázou. V homeostatických podmienkach Nrf2 ovplyvňuje mitochondriálny membránový potenciál, oxidáciu mastných kyselín, dostupnosť substrátov (NADH a FADH2/sukcinát) pre dýchanie a syntézu ATP. V podmienkach stresu alebo stimulácie rastovým faktorom aktivácia Nrf2 pôsobí proti zvýšenej produkcii reaktívnych foriem kyslíka v mitochondriách prostredníctvom transkripčnej upregulácie uncoupling proteínu 3 a ovplyvňuje mitochondriálnu biogenézu udržiavaním hladín jadrového respiračného faktora 1 a receptora aktivovaného peroxizómovým proliferátorom? koaktivátorom 1p, ako aj podporou biosyntézy purínových nukleotidov. Farmakologické aktivátory Nrf2, ako je prirodzene sa vyskytujúci izotiokyanát sulforafan, inhibujú oxidantom sprostredkované otvorenie pórov prechodu mitochondriálnej permeability a opuch mitochondrií. Je zvláštne, že sa zistilo, že syntetická 1,4-difenyl-1,2,3-triazolová zlúčenina, pôvodne navrhnutá ako aktivátor Nrf2, podporuje mitofágiu, čím prispieva k celkovej mitochondriálnej homeostáze. Nrf2 je teda popredným hráčom pri podpore štrukturálnej a funkčnej integrity mitochondrií a táto úloha je obzvlášť kľúčová v podmienkach stresu.
Nrf2 podporuje štrukturálnu a funkčnú integritu mitochondrií.
Aktivátory Nrf2 majú priaznivé účinky, keď je ohrozená mitochondriálna funkcia.
úvod
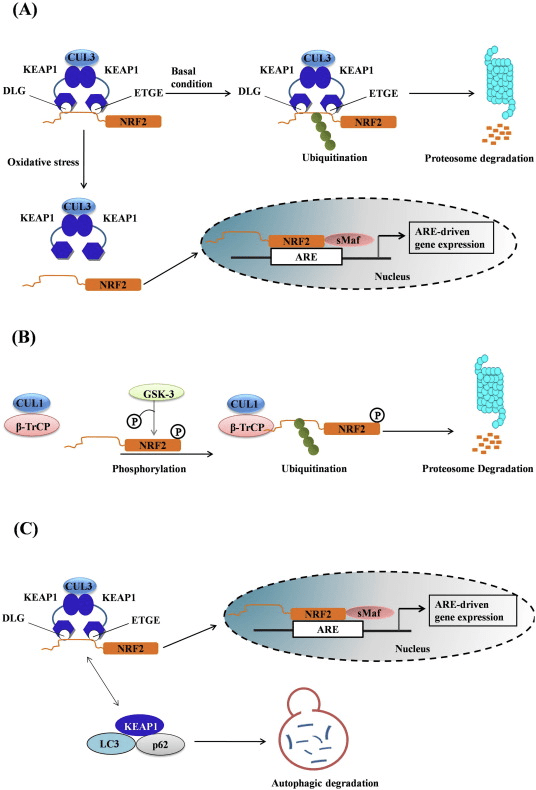
Transkripčný faktor NF-E2 faktor 45 súvisiaci s p2 (Nrf2; názov génu NFE2L2) reguluje expresiu sietí génov kódujúcich proteíny s rôznymi cytoprotektívnymi aktivitami. Samotný Nrf2 je kontrolovaný predovšetkým na úrovni proteínovej stability. Za bazálnych podmienok je Nrf2 proteín s krátkou životnosťou, ktorý podlieha kontinuálnej ubikvitinácii a proteazomálnej degradácii. Existujú tri známe systémy ubikvitín ligázy, ktoré prispievajú k degradácii Nrf2. Historicky prvým negatívnym regulátorom Nrf2, ktorý bol objavený, bol Kelch-like ECH-asociovaný proteín 1 (Keap1) [1], substrátový adaptorový proteín pre ubikvitín ligázu Cullin 3 (Cul3)/Rbx1 [2], [3], [ 4]. Keap1 využíva vysoko účinný cyklický mechanizmus na zacielenie Nrf2 na ubikvitináciu a proteazomálnu degradáciu, počas ktorej sa Keap1 kontinuálne regeneruje, čo umožňuje priebeh cyklu (obr. 1A) [5]. Nrf2 je tiež vystavený degradácii sprostredkovanej ubikvitín ligázou na báze glykogénsyntázy (GSK)3/a-TrCP-dependentnej Cul1 [6], [7]. Najnovšie sa uvádza, že v podmienkach stresu endoplazmatického retikula je Nrf2 ubikvitinovaný a degradovaný v procese sprostredkovanom E3 ubikvitín ligázou Hrd1 [8].
Obrázok 1 Model cyklickej sekvenčnej väzby a regenerácie pre Keap1-sprostredkovanú degradáciu Nrf2. (A) Nrf2 sa viaže postupne na voľný dimér Keap1: najprv prostredníctvom svojej vysokoafinitnej väzbovej domény ETGE (červené tyčinky) a potom prostredníctvom svojej väzbovej domény DLG (čierne tyčinky) s nízkou afinitou. V tejto konformácii proteínového komplexu Nrf2 podlieha ubikvitinácii a je zameraný na proteazomálnu degradáciu. Voľný Keap1 je regenerovaný a schopný viazať sa na novo preložený Nrf2 a cyklus začína znova. (B) Induktory (biele kosoštvorce) reagujú so senzorovými cysteínmi Keap1 (modré tyčinky), čo vedie ku konformačnej zmene a zhoršenej aktivite substrátového adaptéra. Voľný Keap1 sa neregeneruje a novo syntetizovaný Nrf2 sa hromadí a premiestňuje do jadra.
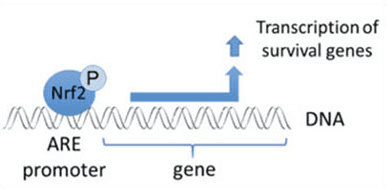
Okrem toho, že Keap1 slúži ako adaptérový proteín substrátu ubikvitín ligázy, je tiež senzorom pre širokú škálu nízkomolekulových aktivátorov Nrf2 (nazývaných induktory) [9]. Induktory blokujú cyklus degradácie Nrf1 sprostredkovanej Keap2 chemickou modifikáciou špecifických cysteínových zvyškov v Keap1 [10], [11] alebo priamym narušením väzbového rozhrania Keap1:Nrf2 [12], [13]. V dôsledku toho nie je Nrf2 degradovaný a transkripčný faktor sa hromadí a translokuje do jadra (obr. 1B), kde tvorí heterodimér s malým proteínom Maf; viaže sa na prvky antioxidačnej odozvy, upstream regulačné oblasti svojich cieľových génov; a iniciuje transkripciu [14], [15], [16]. Batéria cieľov Nrf2 obsahuje proteíny s rôznymi cytoprotektívnymi funkciami, vrátane enzýmov metabolizmu xenobiotík, proteínov s antioxidačnými a protizápalovými funkciami a proteazomálnych podjednotiek, ako aj proteínov, ktoré regulujú bunkovú redoxnú homeostázu a podieľajú sa na intermediárnom metabolizme.
Nrf2: Hlavný regulátor bunkovej redoxnej homeostázy
Funkcia Nrf2 ako hlavného regulátora bunkovej redoxnej homeostázy je široko uznávaná. Génová expresia katalytickej aj regulačnej podjednotky β-glutamylcysteín ligázy, enzýmu katalyzujúceho rýchlosť obmedzujúci krok v biosyntéze redukovaného glutatiónu (GSH), je priamo regulovaná Nrf2 [17]. Podjednotka xCT systému xc-, ktorá importuje cystín do buniek, je tiež priamym transkripčným cieľom Nrf2 [18]. V bunke cystín prechádza konverziou na cysteín, prekurzor pre biosyntézu GSH. Okrem svojej úlohy v biosyntéze GSH, Nrf2 poskytuje prostriedky na udržanie glutatiónu v redukovanom stave koordinovanou transkripčnou reguláciou glutatiónreduktázy 1 [19], [20], ktorá redukuje oxidovaný glutatión na GSH pomocou redukčných ekvivalentov z NADPH. . Požadovaný NADPH zabezpečujú štyri hlavné enzýmy generujúce NADPH, jablčný enzým 1 (ME1), izocitrátdehydrogenáza 1 (IDH1), glukóza-6-fosfátdehydrogenáza (G6PD) a 6-fosfoglukonátdehydrogenáza (PGD), z ktorých všetky sú transkripčne regulované čiastočne Nrf2 (obr. 2) [21], [22], [23], [24]. Je zvláštne, že Nrf2 tiež reguluje indukovateľnú génovú expresiu cytosolických, mikrozomálnych a mitochondriálnych foriem aldehyddehydrogenázy [25], ktoré využívajú NAD(P)+ ako kofaktor, čím vzniká NAD(P)H. Hladiny NADPH a pomer NADPH/NADP+ sú skutočne nižšie v embryonálnych fibroblastoch izolovaných z myší s knockoutom Nrf2 (Nrf2-KO) v porovnaní s bunkami z ich náprotivkov divokého typu (WT) a hladiny NADPH sa znižujú po knockdowne Nrf2 v r. rakovinové bunkové línie s konštitutívne aktívnym Nrf2 [26]. Ako sa očakávalo, hladiny GSH sú nižšie v bunkách, v ktorých bol Nrf2 narušený; naopak, aktivácia Nrf2 genetickými alebo farmakologickými prostriedkami vedie k upregulácii GSH [27], [28], [29]. Dôležité je, že Nrf2 tiež reguluje génovú expresiu tioredoxínu [30], [31], [32], tioredoxín reduktázy 1 [28], [29], [32], [33] a sulfiredoxínu [34], ktoré sú esenciálne na redukciu oxidovaných proteínových tiolov.
Obrázok 2 Úloha Nrf2 v metabolizme rýchlo proliferujúcich buniek. Nrf2 je pozitívny regulátor génov kódujúcich enzýmy v oxidačnom ramene [tj glukóza-6-fosfát dehydrogenáza (G6PD) a 6-fosfoglukonát dehydrogenáza (PGD)] a neoxidačnom ramene [tj transaldoláza 1 (TALDO1) a transketoláza ( TKT)] pentózofosfátovej dráhy. G6PD a PGD generujú NADPH. Nrf2 tiež reguluje génovú expresiu ďalších dvoch enzýmov generujúcich NADPH, jablčného enzýmu 1 (ME1) a izocitrátdehydrogenázy 1 (IDH1). Génová expresia fosforibozylpyrofosfátamidotransferázy (PPAT), ktorá katalyzuje vstup do de novo purínovej biosyntetickej dráhy, je tiež pozitívne regulovaná Nrf2, rovnako ako expresia metyléntetrahydrofolátdehydrogenázy 2 (MTHFD2), mitochondriálneho enzýmu s kritickou úlohou v poskytovanie jednouhlíkových jednotiek pre de novo biosyntézu purínov. Pyruvátkináza (PK) je negatívne regulovaná Nrf2 a očakáva sa, že uprednostňuje hromadenie glykolytických medziproduktov a spolu s G6PD metabolitový kanál cez pentózofosfátovú dráhu a syntézu nukleových kyselín, aminokyselín a fosfolipidov. Nrf2 negatívne reguluje génovú expresiu ATP-citrát lyázy (CL), čo môže zvýšiť dostupnosť citrátu pre mitochondriálne využitie alebo (prostredníctvom izocitrátu) pre IDH1. Červená a modrá označujú pozitívnu a negatívnu reguláciu. Mitochondrie sú zobrazené sivou farbou. Skratky metabolitov: G-6-P, glukóza 6-fosfát; F-6-P, fruktóza-6-fosfát; F-1,6-BP, fruktóza 1,6-bisfosfát; GA-3-P, glyceraldehyd-3-fosfát; 3-PG, 3-fosfoglycerát; PEP, fosfoenolpyruvát; 6-P-Gl, 6-fosfoglukonolaktón; 6-PG, 6-fosfoglukonát; R-5-P, ribulóza 5-fosfát; PRPP, 5-fosforibozyl-a-1-pyrofosfát; THF, tetrahydrofolát; IMP, inozínmonofosfát; AMP, adenozínmonofosfát; GMP, guanozínmonofosfát.
Vzhľadom na kľúčovú úlohu Nrf2 ako hlavného regulátora bunkovej redoxnej homeostázy nie je prekvapujúce, že v porovnaní s bunkami WT sú hladiny reaktívnych foriem kyslíka (ROS) vyššie v bunkách, v ktorých bol Nrf2 narušený (Nrf2-KO) [35]. Tento rozdiel je obzvlášť markantný pri napadnutí látkami, ktoré spôsobujú oxidačný stres. Navyše bunky s deficitom Nrf2 sú oveľa citlivejšie na toxicitu oxidantov rôznych typov a nemôžu byť chránené induktormi Nrf2, ktoré za rovnakých podmienok poskytujú účinnú a dlhotrvajúcu ochranu bunkám WT [29], [36] [37]. Okrem celkovej bunkovej redoxnej homeostázy je Nrf2 tiež rozhodujúci pre udržanie mitochondriálnej redoxnej homeostázy. V porovnaní s WT je teda celkový mitochondriálny súbor NADH významne zvýšený v Keap1-KO a dramaticky znížený v bunkách Nrf2-KO [35].
Pomocou zobrazenia živých buniek sme nedávno monitorovali rýchlosti produkcie ROS v primárnych glioneuronálnych kokultúrach a rezoch mozgového tkaniva izolovaných z myší WT, Nrf2-KO alebo Keap1-knockdown (Keap1-KD) [38]. Ako sa očakávalo, rýchlosť produkcie ROS bola rýchlejšia v bunkách a tkanivách Nrf2-KO v porovnaní s ich náprotivkami WT. Urobili sme však neočakávané pozorovanie, že v porovnaní s WT majú bunky Keap1-KD tiež vyššiu mieru produkcie ROS, hoci veľkosť rozdielu medzi genotypmi WT a Keap1-KD bola menšia ako medzi WT a Nrf2-KO. . Potom sme analyzovali hladiny mRNA NOX2 a NOX4, katalytických podjednotiek dvoch izoforiem NADPH oxidázy (NOX), ktoré sa podieľajú na patológii mozgu, a zistili sme, že NOX2 sa dramaticky zvyšuje v podmienkach nedostatku Nrf2, zatiaľ čo NOX4 je upregulovaný, keď Nrf2 sa konštitutívne aktivuje, aj keď v menšej miere. Kvantitatívne je veľkosť upregulácie v bunkách a tkanivách z mutantných myší paralelná so zodpovedajúcim zvýšením produkcie ROS [38]. Je zaujímavé, že Nrf2 nielen reguluje NADPH oxidázu, ale aj ROS produkovaný NADPH oxidázou môže aktivovať Nrf2, ako je ukázané v pľúcnych epiteliálnych bunkách a kardiomyocytoch [39], [40]. Okrem toho veľmi nedávna štúdia preukázala, že aktivácia Nrf2 závislá od NADPH oxidázy predstavuje dôležitý endogénny mechanizmus ochrany pred mitochondriálnym poškodením a bunkovou smrťou v srdci počas chronického tlakového preťaženia [41].
Okrem katalytickej aktivity NADPH oxidázy je ďalším hlavným intracelulárnym zdrojom ROS mitochondriálne dýchanie. Použitím mitochondriálnej špecifickej sondy MitoSOX sme skúmali príspevok ROS mitochondriálneho pôvodu k celkovej produkcii ROS v primárnych glioneuronálnych izolovaných kokultúrach z myší WT, Nrf2-KO alebo Keap1-KD [38]. Ako sa očakávalo, bunky Nrf2-KO mali vyššiu mieru mitochondriálnej produkcie ROS ako WT. V súlade so zisteniami o celkovej produkcii ROS boli miery produkcie mitochondriálnych ROS v Keap1-KD tiež vyššie v porovnaní s bunkami WT. Dôležité je, že blokovanie komplexu I s rotenónom spôsobilo dramatický nárast mitochondriálnej produkcie ROS v bunkách WT aj Keap1-KD, ale nemalo žiadny účinok v bunkách Nrf2-KO. Na rozdiel od očakávaného zvýšenia produkcie mitochondriálnych ROS v bunkách WT po pridaní pyruvátu (na zvýšenie dostupnosti NADH, zvýšenie potenciálu mitochondriálnej membrány a normalizáciu dýchania) sa produkcia ROS v bunkách Nrf2-KO znížila. Spoločne tieto zistenia silne naznačujú, že v neprítomnosti Nrf2: (i) je narušená aktivita komplexu I, (ii) narušená aktivita komplexu I je spôsobená obmedzením substrátov a (iii) narušená aktivita komplexu I je jedným z hlavných dôvodov zvýšenej mitochondriálnej produkcie ROS, pravdepodobne v dôsledku reverzného toku elektrónov z komplexu II.
Nrf2 ovplyvňuje mitochondriálny membránový potenciál a dýchanie
Mitochondriálny membránový potenciál (??m) je univerzálnym indikátorom mitochondriálneho zdravia a metabolického stavu bunky. V zdravej bunke je µm udržiavaný mitochondriálnym dýchacím reťazcom. Je zaujímavé, že stabilné izotopové značenie aminokyselinami v proteomickej štúdii založenej na kultúre v netumorogénnej bunkovej línii ľudského prsného epitelu MCF10A s negatívnym estrogénovým receptorom ukázalo, že zložka mitochondriálneho elektrónového transportného reťazca NDUFA4 je upregulovaná farmakologickou aktiváciou (sulforafanom) Nrf2, zatiaľ čo genetická upregulácia Nrf2 (knockdown Keap1) vedie k downregulácii podjednotiek cytochróm c oxidázy COX2 a COX4I1 [42]. Štúdia pečeňového proteómu pomocou dvojrozmernej gélovej elektroforézy a hmotnostnej spektrometrie s laserovou desorpciou/ionizáciou za pomoci matrice zistila, že Nrf2 reguluje expresiu podjednotky ATP syntázy ? [43]. Okrem toho sa uvádza, že mitochondriálny proteín DJ-1, ktorý hrá úlohu pri udržiavaní aktivity komplexu I [44], stabilizuje Nrf2 [45], [46], hoci neuroprotektívne účinky farmakologickej alebo genetickej aktivácie z Nrf2 sú nezávislé od DJ-1 [47]. Dôsledky týchto pozorovaní na mitochondriálnu funkciu však neboli skúmané.
V súlade so zhoršenou aktivitou komplexu I v podmienkach deficitu Nrf2 je bazálny ??m nižší v Nrf2-KO myších embryonálnych fibroblastoch (MEF) a kultivovaných primárnych glioneuronálnych bunkách v porovnaní s ich WT náprotivkami (obr. 3, vložka) [35]. Na rozdiel od toho, bazálny ??m je vyšší, keď je Nrf2 geneticky konštitutívne upregulovaný (knockdown alebo knockout Keap1). Tieto rozdiely v µm medzi genotypmi naznačujú, že dýchanie je ovplyvnené aktivitou Nrf2. Vyhodnotenie spotreby kyslíka v bazálnom stave skutočne odhalilo, že v porovnaní s WT je spotreba kyslíka nižšia v Nrf2-KO a Keap1-KO MEF o ~50 a ~35%.
Obrázok 3 Navrhovaný mechanizmus narušenej mitochondriálnej funkcie v podmienkach nedostatku Nrf2. (1) Znížené hladiny ME1, IDH1, G6PD a PGD vedú k nižším hladinám NADPH. (2) Hladiny GSH sú tiež nízke. (3) Nízka aktivita ME1 môže znížiť množstvo pyruvátu vstupujúceho do mitochondrií. (4) Tvorba NADH je pomalšia, čo vedie k zhoršenej aktivite komplexu I a zvýšenej mitochondriálnej produkcii ROS. (5) Zníženie FAD na FADH2 v mitochondriálnych proteínoch je tiež znížené, čím sa znižuje tok elektrónov z FADH2 do UbQ a do komplexu III. (6) Pomalšia tvorba UbQH2 môže znížiť enzýmovú aktivitu sukcinátdehydrogenázy. (7) Zvýšené hladiny ROS môžu ďalej inhibovať aktivitu komplexu II. (8) Nižšia účinnosť oxidácie mastných kyselín prispieva k zníženej dostupnosti substrátu pre mitochondriálne dýchanie. (9) Glykolýza je posilnená ako kompenzačný mechanizmus pre zníženú produkciu ATP pri oxidatívnej fosforylácii. (10) ATP syntáza pracuje opačne, aby udržala ?m. Červená a modrá označujú upreguláciu a downreguláciu. Políčka označujú dostupnosť experimentálnych dôkazov. Vložka ukazuje obrázky mitochondrií kortikálnych astrocytov WT a Nrf2-KO vizualizované potenciometrickou fluorescenčnou sondou tetrametylrodamín metylester (TMRM; 25 nM). Mierka, 20 �m.
Tieto rozdiely v µm a dýchaní medzi genotypmi sa odrážajú v miere využitia substrátov na mitochondriálne dýchanie. Aplikácia substrátov pre cyklus trikarboxylových kyselín (TCA) (jablčnan/pyruvát, ktoré následne zvyšujú produkciu komplexu I, substrát NADH) alebo metylsukcinátu, substrátu pre komplex II, spôsobuje postupné zvyšovanie ??m v oboch WT a Keap1-KD neuróny, ale rýchlosť nárastu je vyššia v Keap1-KD bunkách. Ešte dôležitejšie je, že tvary odozvy na tieto substráty cyklu TCA sa medzi týmito dvoma genotypmi líšia, pričom rýchly nárast µm v bunkách Keap1-KD po pridaní substrátu je nasledovaný skôr rýchlym poklesom než plató, čo naznačuje nezvyčajne rýchla spotreba substrátu. Tieto zistenia sú v úzkej zhode s oveľa nižšími (o 50–70 %) hladinami malátu, pyruvátu a sukcinátu, ktoré boli pozorované po 1-hodinovom pulze [U-13C6]glukózy v Keap1-KO v porovnaní s WT MEF bunky [24]. V neurónoch Nrf2-KO je iba pyruvát schopný zvýšiť ??m, zatiaľ čo malát a metylsukcinát spôsobujú miernu depolarizáciu. Účinok Nrf2 na produkciu mitochondriálneho substrátu sa zdá byť hlavným mechanizmom, ktorým Nrf2 ovplyvňuje mitochondriálnu funkciu. Mitochondriálny redox index NADH (rovnováha medzi spotrebou NADH komplexom I a produkciou NADPH v cykle TCA) je výrazne nižší v bunkách Nrf2-KO v porovnaní s ich náprotivkami WT a navyše rýchlosti regenerácie poolov NADH a FADH2 po inhibícii komplexu IV (pomocou NaCN) sú v mutantných bunkách pomalšie.
V mitochondriách izolovaných z myšieho mozgu a pečene suplementácia substrátov pre komplex I alebo pre komplex II zvyšuje rýchlosť spotreby kyslíka výraznejšie, keď je aktivovaný Nrf2 a menej efektívne, keď je Nrf2 narušený [35]. Malát teda indukuje vyššiu mieru spotreby kyslíka v Keap1-KD v porovnaní s WT, ale jeho účinok je slabší v mitochondriách Nrf2-KO. Podobne v prítomnosti rotenónu (keď je komplex I inhibovaný) sukcinát aktivuje spotrebu kyslíka vo väčšej miere v Keap1-KD v porovnaní s WT, zatiaľ čo odpoveď v mitochondriách Nrf2-KO je znížená. Okrem toho sú primárne neurónové kultúry a myši Nrf2-KO citlivejšie na toxicitu inhibítorov komplexu II 3-nitropropiónovej kyseliny a malonátu, zatiaľ čo intrastriatálna transplantácia astrocytov s nadmernou expresiou Nrf2 je ochranná [48], [49]. Podobne aj myši Nrf2-KO sú citlivejšie na, zatiaľ čo genetická alebo farmakologická aktivácia Nrf2 má ochranné účinky proti neurotoxicite spôsobenej inhibítorom komplexu I 1-metyl-4-fenylpyridínium ión v 1-metyl-4-fenyl-1,2,3,6, 49-tetrahydropyridínový zvierací model Parkinsonovej choroby [50], [51], [52], [53], [54], [55], [56], [57], [58], [59], [60], [61], [XNUMX].
Pomer kontroly dýchania (RCR), pomer stavu 3 (stimulované ADP) k dýchaniu v stave 4 (bez prítomnosti ADP), je znížený v neprítomnosti Nrf2, ale RCR je podobný medzi Keap1-KD a mitochondriami WT [35 ]. Keďže RCR je indikáciou stupňa spojenia aktivity mitochondriálneho dýchacieho reťazca s oxidačnou fosforyláciou, toto zistenie naznačuje, že vyššia rýchlosť dýchania v mitochondriách Keap1-KD nie je spôsobená odpojením oxidatívnej fosforylácie. Ďalej to naznačuje, že oxidačná fosforylácia je účinnejšia, keď je aktivovaný Nrf2. Vyššia rýchlosť dýchania v mitochondriách Keap1-KD je v súlade s vyššími hladinami mitochondriálnej produkcie ROS [38], pretože vyššie rýchlosti dýchania môžu viesť k zvýšenému úniku elektrónov. Avšak v podmienkach oxidačného stresu je zvýšená produkcia ROS neutralizovaná Nrf2-dependentnou transkripčnou upreguláciou uncoupling proteínu 3 (UCP3), ktorá zvyšuje protónovú vodivosť mitochondriálnej vnútornej membrány a následne znižuje produkciu superoxidu [62]. Veľmi nedávno sa ukázalo, že produkt peroxidácie lipidov 4-hydroxy-2-nonenal sprostredkuje Nrf2-dependentnú upreguláciu UCP3 v kardiomyocytoch; to môže byť obzvlášť dôležité pre ochranu v podmienkach oxidačného stresu, ako sú tie počas ischemickej reperfúzie [63].
Nrf2 ovplyvňuje účinnosť oxidačnej fosforylácie a syntézu ATP
V súlade s účinkom Nrf2 na dýchanie v mitochondriách mozgu a pečene vedie nedostatok Nrf2 k zníženiu účinnosti oxidačnej fosforylácie (ako sa odhaduje pomerom ADP ku kyslíku, ktorý sa spotrebuje na syntézu ATP), zatiaľ čo aktivácia Nrf2 (Keap1 -KD) má opačný účinok [35]. V porovnaní s WT sú hladiny ATP významne vyššie v bunkách s konštitutívnou upreguláciou Nrf2 a nižšie, keď je Nrf2 zrazený [64] alebo narušený [35]. Okrem toho použitie inhibítorov oxidatívnej fosforylácie (oligomycín) alebo glykolýzy (kyselina jódoctová) odhalilo, že Nrf2 mení spôsob, akým bunky produkujú ATP. V neurónoch WT teda oligomycín spôsobuje úplný pokles ATP a kyselina jódoctová už nemá žiadny ďalší účinok. Je pozoruhodné, že v bunkách Nrf2-KO oligomycín zvyšuje hladiny ATP, ktoré sa potom pomaly, ale úplne vyčerpávajú kyselinou jódoctovou, čo naznačuje, že v neprítomnosti Nrf2 je hlavným zdrojom produkcie ATP glykolýza a nie oxidačná fosforylácia. Je zaujímavé, že napriek zvýšenej účinnosti oxidačnej fosforylácie v bunkách Keap1-KD má pridanie oligomycínu za následok ~80% zníženie hladín ATP a kyselina jódoctová spôsobuje ďalší ~20% pokles. Deficit Nrf2 alebo jeho konštitutívna aktivácia teda znižuje príspevok oxidatívnej fosforylácie a zvyšuje príspevok glykolýzy k syntéze ATP. Tento účinok je obzvlášť výrazný, keď chýba Nrf2 a je v súlade so závislosťou ??m od prítomnosti glukózy v médiu [35] a zvýšených hladín glykolytických medziproduktov (G-6-P, F-6-P , dihydroxyacetón fosfát, pyruvát a laktát) po knockdown Nrf2 [24].
Zvýšenie hladín ATP po inhibícii F1F0-ATPázy oligomycínom naznačuje, že v neprítomnosti Nrf2 funguje F1F0-ATPáza ako ATPáza a nie ako ATP syntáza, tj funguje opačne. Takéto zvrátenie aktivity s najväčšou pravdepodobnosťou odráža potrebu pumpovať protóny cez vnútornú mitochondriálnu membránu v snahe udržať µm, čo je rozhodujúce pre funkčnú integritu tejto organely. O zvrátení funkcie F1F0-ATPázy svedčí aj pozorovaná mitochondriálna depolarizácia po podaní oligomycínu bunkám Nrf2-KO, čo je v ostrom kontraste s hyperpolarizáciou vyskytujúcou sa u ich náprotivkov s deficitom WT alebo Keap1 [35]. Celkovo sa zdá, že za podmienok nedostatku Nrf2 sa ATP produkuje primárne v glykolýze a tento ATP je potom čiastočne využívaný F1F0-ATPázou na udržanie ?m.
Nrf2 zvyšuje mitochondriálnu oxidáciu mastných kyselín
Účinok nedostatku Nrf2 na ??m je obzvlášť výrazný, keď sú bunky inkubované v médiu bez glukózy, a ??m je v Nrf50-KO ~2 % nižší v porovnaní s bunkami WT [35]. V podmienkach nedostatku glukózy je oxidácia mitochondriálnych mastných kyselín (FAO) hlavným poskytovateľom substrátov pre dýchanie a oxidačnú fosforyláciu, čo naznačuje, že Nrf2 môže ovplyvniť FAO. V skutočnosti je účinnosť FAO pre nasýtenú mastnú kyselinu palmitovú s dlhým reťazcom (C16:0) a kyselinu hexánovú s krátkym reťazcom (C6:0) vyššia v Keap1-KO MEF a izolovaných srdcových a pečeňových mitochondriách ako v ich náprotivky WT, zatiaľ čo v bunkách Nrf2-KO a mitochondriách je nižšia [65]. Tieto účinky sú tiež vysoko relevantné pre ľudí: skutočne sa uvádza, že metabolické zmeny naznačujúce lepšiu integráciu FAO s aktivitou cyklu TCA sa vyskytujú v intervenčných štúdiách u ľudí s diétami bohatými na glukorafanín, prekurzor klasického aktivátora Nrf2 sulforafanu [ 66].
Počas prvého kroku mitochondriálnej FAO odchádza pro-R vodík z a-uhlíka ako hydrid, ktorý redukuje FAD kofaktor na FADH2, ktorý následne prenáša elektróny na ubichinón (UbQ) v dýchacom reťazci, čo v konečnom dôsledku prispieva k produkcii ATP. . Zatiaľ čo stimulácia FAO palmitoylkarnitínom v neprítomnosti glukózy spôsobuje očakávané zvýšenie hladín ATP v bunkách WT a Keap1-KO, pričom nárast ATP je rýchlejší v bunkách Keap1-KO, identická liečba neprodukuje žiadne zmeny ATP v Nrf2-KO MEF [65]. Tento experiment ukazuje, že v neprítomnosti Nrf2 je FAO potlačená a okrem toho implikuje supresiu FAO ako jeden z dôvodov nižších hladín ATP v podmienkach nedostatku Nrf2 [35], [64].
Je pozoruhodné, že ľudské 293 T bunky, v ktorých bol Nrf2 umlčaný, majú nižšiu expresiu CPT1 a CPT2[67], dvoch izoforiem karnitín palmitoyltransferázy (CPT), enzýmu obmedzujúceho rýchlosť v mitochondriálnej FAO. V zhode sú hladiny mRNA Cpt1 nižšie v pečeni Nrf2-KO v porovnaní s myšami WT [68]. CPT katalyzuje prenos acylovej skupiny mastného acyl-CoA s dlhým reťazcom z koenzýmu A na l-karnitín a umožňuje tak import acylkarnitínu z cytoplazmy do mitochondrií. Hoci to doteraz nebolo preskúmané, je možné, že okrem transkripčných účinkov na expresiu CPT1 môže Nrf2 ovplyvňovať aj funkciu tohto enzýmu riadením hladín jeho hlavného alosterického inhibítora, malonyl-CoA. Je to preto, že mechanizmom, ktorý je v súčasnosti nejasný, Nrf2 negatívne reguluje expresiu stearoyl CoA desaturázy (SCD) [69] a citrát lyázy (CL) [69], [70]. Je zvláštne, že knockout alebo inhibícia SCD vedie k zvýšenej fosforylácii a aktivácii AMP-aktivovanej proteínkinázy (AMPK) [71], [72], [73] a možno špekulovať, že v neprítomnosti Nrf2 budú hladiny SCD sa zvýši, čím sa zníži aktivita AMPK. To by mohlo byť ďalej spojené so zníženými hladinami proteínov AMPK, ktoré boli pozorované v pečeni myší Nrf2-KO [68], čo je zistenie, ktoré je v úzkej zhode so zvýšenými hladinami AMPK, ktoré boli hlásené v pečeni Keap1-KD. myši [74]. Jedným z dôsledkov zníženej aktivity AMPK je zmiernenie jej inhibičnej fosforylácie (na Ser79) acetyl-CoA karboxylázy (ACC) [75], ktorá by mohla byť ďalej transkripčne upregulovaná v neprítomnosti Nrf2, pretože je downregulovaná aktiváciou Nrf2 [70 ]. Vysoká aktivita ACC v kombinácii s upregulovanou expresiou CL, ktorá zvýši produkciu acetyl-CoA, substrátu pre ACC, môže v konečnom dôsledku zvýšiť hladiny produktu ACC, malonyl-CoA. Vysoké hladiny malonyl-CoA budú inhibovať CPT, čím sa zníži transport mastných kyselín do mitochondrií. Nakoniec Nrf2 pozitívne reguluje expresiu CD36 [76], translokázy, ktorá importuje mastné kyseliny cez plazmatické a mitochondriálne membrány. Jedným mechanizmom, ktorým môže Nrf2 ovplyvniť účinnosť mitochondriálnej FAO, je teda regulácia importu mastných kyselín s dlhým reťazcom do mitochondrií.
Okrem priamej transkripčnej regulácie môže Nrf2 tiež meniť účinnosť mitochondriálnej FAO svojimi účinkami na bunkový redoxný metabolizmus. To môže byť obzvlášť dôležité, keď je aktivita Nrf2 nízka alebo chýba, podmienky, ktoré posúvajú bunkový redoxný stav smerom k oxidovanému stavu. V skutočnosti bolo niekoľko enzýmov FAO identifikovaných ako citlivých na redoxné zmeny. Jedným z takýchto enzýmov je acyl-CoA dehydrogenáza s veľmi dlhým reťazcom (VLCAD), ktorá sa podieľa viac ako 80 % na dehydrogenačnej aktivite palmitoyl-CoA v ľudských tkanivách [77]. Zaujímavé je, že Hurd a spol. [78] ukázali, že VLCAD obsahuje cysteínové zvyšky, ktoré významne menia svoj redoxný stav po vystavení izolovaných mitochondrií srdca potkana H2O2. Okrem toho S-nitrozylácia myšieho pečeňového VLCAD na Cys238 zlepšuje katalytickú účinnosť enzýmu [79] a je pravdepodobné, že oxidácia toho istého cysteínu môže mať opačný účinok, čo v konečnom dôsledku znižuje účinnosť mitochondriálnej FAO. Je preto možné, že aj keď sa hladiny expresie VLCAD významne nelíšia vo WT, Nrf2-KO alebo Keap1-KO MEF [65], enzýmová aktivita VLCAD by mohla byť nižšia v neprítomnosti Nrf2 v dôsledku vyšších hladín z ROS.
Na základe všetkých týchto zistení možno navrhnúť, že (obr. 3): v neprítomnosti Nrf2 sú hladiny NADPH nižšie v dôsledku zníženej expresie ME1, IDH1, G6PD a PGD. Hladiny redukovaného glutatiónu sú tiež nižšie v dôsledku zníženej expresie enzýmov, ktoré sa podieľajú na jeho biosyntéze a regenerácii a nižších hladín NADPH, ktoré sú potrebné na premenu oxidovaného na redukovanú formu glutatiónu. Nízka expresia ME1 zníži množstvo pyruvátu vstupujúceho do mitochondrií, pričom hlavným zdrojom pyruvátu sa stane glykolýza. Tvorba NADH je pomalšia, čo vedie k zhoršenej aktivite komplexu I a zvýšenej mitochondriálnej produkcii ROS. Redukcia FAD na FADH2 je tiež pomalšia, aspoň čiastočne v dôsledku menej účinnej oxidácie mastných kyselín, čo ohrozuje tok elektrónov z FADH2 do UbQ a do komplexu III. Keďže UbQH2 je aktivátor sukcinátdehydrogenázy [80], spomalenie jeho tvorby môže znížiť aktivitu enzýmu sukcinátdehydrogenázy. Zvýšené hladiny superoxidu a peroxidu vodíka môžu ďalej inhibovať aktivitu komplexu II [81]. Nižšia účinnosť oxidácie mastných kyselín prispieva k zníženej dostupnosti substrátu pre mitochondriálne dýchanie a produkciu ATP pri oxidatívnej fosforylácii. Ako kompenzačný mechanizmus sa zvyšuje glykolýza. ATP syntáza funguje opačne, ako ATPáza, v snahe udržať ?m.
Nrf2 a mitochondriálna biogenéza
Bolo publikované, že v porovnaní s WT majú pečeň Nrf2-KO myší nižší mitochondriálny obsah (ako je určené pomerom mitochondriálnej a jadrovej DNA); toto je ďalej znížené 24-hodinovým hladovaním u myší WT aj Nrf2-KO; na rozdiel od toho, hoci sa nelíši od WT za normálnych podmienok kŕmenia, obsah mitochondrií u myší s vysokou aktivitou Nrf2 nie je ovplyvnený hladovaním [82]. Je zaujímavé, že suplementácia Nrf2 aktivátorom (R)-a-lipoová kyselina [83], [84], [85] podporuje mitochondriálnu biogenézu v 3T3-L1 adipocytoch [86]. Dve triedy jadrových transkripčných regulátorov hrajú rozhodujúcu úlohu v mitochondriálnej biogenéze. Prvou triedou sú transkripčné faktory, ako sú jadrové respiračné faktory11 a 2, ktoré riadia expresiu génov kódujúcich podjednotky piatich respiračných komplexov, mitochondriálne translačné zložky a enzýmy biosyntézy hemu, ktoré sú lokalizované v mitochondriálnej matrici [88]. Piantadosi a kol. [89] ukázali, že transkripčná upregulácia jadrového respiračného faktora 2 závislá od Nrf1 podporuje mitochondriálnu biogenézu a chráni pred cytotoxicitou kardiotoxického antracyklínového chemoterapeutika doxorubicínu. Na rozdiel od toho Zhang a kol. [82] uviedli, že genetická aktivácia Nrf2 neovplyvňuje bazálnu expresiu mRNA jadrového respiračného faktora 1 v myšacej pečeni.
Druhou triedou jadrových transkripčných regulátorov s kritickými funkciami v mitochondriálnej biogenéze sú transkripčné koaktivátory, ako je receptor aktivovaný peroxizómovým proliferátorom? koaktivátory (PGC)1? a 1p, ktoré interagujú s transkripčnými faktormi, bazálnym transkripčným mechanizmom a mechanizmom zostrihu RNA a enzýmami modifikujúcimi histón [88], [90], [91]. Expresia skupiny koaktivátorov PGC1 je ovplyvnená mnohými environmentálnymi signálmi. Liečba ľudských fibroblastov aktivátorom Nrf2 sulforafanom spôsobuje zvýšenie mitochondriálnej hmoty a indukciu PGC1? a PGC1? [92], hoci potenciálna závislosť od Nrf2 nebola v tejto štúdii skúmaná. Avšak diabetické myši, u ktorých je Nrf2 buď aktivovaný hypomorfným knockdownom génu Keap1 (db/db:Keap1flox/?:Nrf2+/+) alebo narušený (db/db:Keap1flox/?:Nrf2?/?), majú nižšie pečeňové PGC1? úrovne expresie ako kontrolné zvieratá (db/db:Keap1flox/+:Nrf2+/+) [93]. Žiadne rozdiely v hladinách mRNA pre PGC1? sú pozorované v pečeni nediabetických myší, ktoré sú buď WT alebo Nrf2-KO, zatiaľ čo tieto hladiny sú nižšie u zvierat s nadmernou expresiou Nrf2 (Keap1-KD a Keap1-KO špecifický pre pečeň) [82]. Je pozoruhodné, že 24-hodinový pôst zvyšuje hladiny PGC1? mRNA v pečeni myší všetkých genotypov, ale zvýšenie je výrazne väčšie v pečeni Nrf2-KO v porovnaní s myšami s nadmernou expresiou WT alebo Nrf2. V porovnaní s WT myši Nrf2-KO so septickou infekciou alebo akútnym poškodením pľúc v dôsledku infekcie vykazujú zoslabenú transkripčnú upreguláciu jadrového respiračného faktora 1 a PGC1? [94], [95]. Tieto pozorovania spoločne naznačujú, že úloha Nrf2 pri udržiavaní hladín jadrového respiračného faktora 1 a PGC1? je zložitý a najvýraznejší sa stáva v podmienkach stresu.
Okrem expresie génov kódujúcich mitochondriálne proteíny si mitochondriálna biogenéza vyžaduje syntézu nukleotidov. Genetická aktivácia Nrf2 zvyšuje biosyntézu purínov zvýšením regulácie pentózofosfátovej dráhy a metabolizmu folátu a glutamínu, najmä v rýchlo proliferujúcich bunkách (obr. 2) [24]. Analýza transkriptómu mutantnej Drosophila s deficitom mitochondriálnej serín/treonínovej proteínkinázy PTEN-indukovanej predpokladanej kinázy 1 (PINK1) ukázala, že mitochondriálna dysfunkcia vedie k transkripčnej upregulácii génov ovplyvňujúcich metabolizmus nukleotidov [96], čo naznačuje, že zvýšená biosyntéza nukleotidov predstavuje mechanizmus ochrany pred neurotoxickými následkami nedostatku PINK1. Nrf2 reguluje expresiu fosforibozylpyrofosfátamidotransferázy (PPAT), ktorá katalyzuje vstup do de novo biosyntetickej dráhy purínového nukleotidu, a mitochondriálnej metyléntetrahydrofolátdehydrogenázy 2 (MTHFD2) (obr. 2). Posledne menovaný je bifunkčný enzým s dehydrogenázovou a cyklohydrolázovou aktivitou, ktorý je rozhodujúci pri poskytovaní glycínu aj formiátu ako zdrojov jednouhlíkových jednotiek na biosyntézu purínov v rýchlo rastúcich bunkách [97]. Je preto pravdepodobné, že aktivácia Nrf2 môže byť ochranná a môže zvrátiť mitochondriálnu dysfunkciu pri deficite PINK1. Farmakologická aktivácia Nrf2 sulforafanom alebo triterpenoidom RTA-408 skutočne obnovuje µm a chráni bunky s deficitom PINK1 pred dopamínovou toxicitou [98]. Aj keď sa zdá, že základné mechanizmy sú zložité, tieto zistenia spolu naznačujú, že aktivita Nrf2 môže ovplyvniť mitochondriálnu biogenézu ovplyvnením hladín expresie kritických transkripčných faktorov a koaktivátorov, ako aj zvýšením biosyntézy nukleotidov.
Nrf2 a mitochondriálna integrita
Hoci priamy dôkaz nie je vždy dostupný, existujú silné náznaky, že Nrf2 je dôležitý pre mitochondriálnu integritu, najmä v podmienkach oxidačného stresu. Mitochondrie izolované z mozgu a pečene potkanov, ktorým bola podaná jednorazová dávka aktivátora Nrf2 sulforafanu, sú odolné voči otvoreniu mitochondriálneho priepustného prechodového póru (mPTP) spôsobeného oxidantom terc-butylhydroperoxid [99], [100]. Nedávno sa zistilo, že mPTP, komplex, ktorý umožňuje, aby sa vnútorná membrána mitochondrií stala priepustnou pre molekuly s hmotnosťou do 1500 Da, sa tvorí z dimérov F0F1-ATP syntázy [101]. Sulfirafánom sprostredkovaná rezistencia na otvorenie mPTP koreluje so zvýšenou antioxidačnou obranou a hladiny mitochondriálneho GSH, glutatiónperoxidázy 1, jablčného enzýmu 3 a tioredoxínu 2 sú všetky upregulované v mitochondriálnych frakciách izolovaných zo zvierat liečených sulforafanom [100].
Poškodenie mitochondriálneho proteínu a zhoršenie dýchania spôsobené produktom elektrofilnej peroxidácie lipidov 4-hydroxy-2-nonenalom sú oslabené v mitochondriách izolovaných z mozgovej kôry myší liečených sulforafanom [102]. V renálnych epiteliálnych bunkách potkanov a v obličkách sulforafan chráni pred toxicitou vyvolanou cisplatinou a gentamicínom a stratou µm[103], [104]. Ochrana pred skupinou oxidantov (superoxid, peroxid vodíka, peroxydusitan) a elektrofilmi (4-hydroxy-2-nonenal a akroleín) a zvýšenie mitochondriálnej antioxidačnej obrany bola tiež pozorovaná pri liečbe buniek hladkého svalstva potkanej aorty sulforafanom [105 ]. V modeli akútneho poškodenia obličiek vyvolaného kontrastom sa nedávno ukázalo, že predkondicionovanie ischemickej končatiny má ochranné účinky, vrátane inhibície otvorenia mPTP a mitochondriálneho opuchu, aktiváciou Nrf2 v dôsledku inhibície GSK3? [106].
Mitofágia, proces, ktorým sú dysfunkčné mitochondrie selektívne pohltené autofagozómami a dodávané do lyzozómov, aby boli degradované a recyklované bunkou, je nevyhnutný pre mitochondriálnu homeostázu [107], [108]. Zatiaľ čo nebol stanovený žiadny príčinný vzťah medzi Nrf2 a mitofágiou, existujú dôkazy, že transkripčný faktor môže byť dôležitý pri kontrole mitochondriálnej kvality tým, že hrá úlohu v mitofágii. To môže byť obzvlášť výrazné v podmienkach oxidačného stresu. V modeli sepsy je teda zvýšenie hladín autofagozómového markera MAP1 ľahkého reťazca 3-II (LC3-II) a cargo proteínu p62 po 24 hodinách po infekcii u Nrf2-KO v porovnaní s myšami WT potlačené [109]. . Nedávno bol objavený induktor mitofágie s malou molekulou (nazývaný induktor mitofágie sprostredkovaný p62, PMI); táto 1,4-difenyl-1,2,3-triazolová zlúčenina bola pôvodne navrhnutá ako aktivátor Nrf2, ktorý narúša interakciu transkripčného faktora s Keap1 [110]. Podobne ako bunky, v ktorých je Nrf2 geneticky upregulovaný (Keap1-KD alebo Keap1-KO), bunky vystavené PMI majú vyšší pokojový ?m. Dôležité je, že zvýšenie mitochondriálnej lokalizácie LC3, ktoré sa pozoruje po ošetrení buniek WT PMI, sa nevyskytuje v bunkách Nrf2-KO, čo naznačuje zapojenie Nrf2.
Nakoniec ultraštrukturálna analýza pečeňových rezov odhalila prítomnosť opuchnutých mitochondrií so zníženou kristou a narušenými membránami v hepatocytoch myší Nrf2-KO, ale nie WT, ktoré boli 24 týždňov kŕmené stravou s vysokým obsahom tukov; najmä tieto pečene vykazujú jasný dôkaz oxidačného stresu a zápalu [68]. Možno konštatovať, že Nrf2 má rozhodujúcu úlohu pri udržiavaní mitochondriálnej integrity v podmienkach oxidačného a zápalového stresu.
Sulforaphane a jeho účinky na rakovinu, úmrtnosť, starnutie, mozog a správanie, srdcové choroby a ďalšie
Izotiokyanáty sú niektoré z najdôležitejších rastlinných zlúčenín, ktoré môžete získať vo svojej strave. V tomto videu pre nich robím najkomplexnejší prípad, aký kedy bol vyrobený. Krátka doba pozornosti? Preskočte na svoju obľúbenú tému kliknutím na jeden z časových bodov nižšie. Úplná časová os nižšie.
Kľúčové sekcie:
00:01:14 – Rakovina a úmrtnosť
00:19:04 – Starnutie
00:26:30 – Mozog a správanie
00:38:06 – Záverečná rekapitulácia
00:40:27 – Dávka
Úplná časová os:
00:00:34 – Predstavenie sulforafanu, hlavné zameranie videa.
00:01:14 – Spotreba krížovej zeleniny a zníženie úmrtnosti zo všetkých príčin.
00:02:12 – Riziko rakoviny prostaty.
00:02:23 – Riziko rakoviny močového mechúra.
00:02:34 – Riziko rakoviny pľúc u fajčiarov.
00:02:48 – Riziko rakoviny prsníka.
00:03:13 – Hypotetické: čo ak už máte rakovinu? (intervenčné)
00:03:35 – Pravdepodobný mechanizmus, ktorý riadi asociatívne údaje o rakovine a úmrtnosti.
00:04:38 – Sulforafán a rakovina.
00:05:32 – Dôkazy na zvieratách ukazujúce silný účinok extraktu z výhonkov brokolice na vývoj nádoru močového mechúra u potkanov.
00:06:06 – Účinok priamej suplementácie sulforafanu u pacientov s rakovinou prostaty.
00:07:09 – Bioakumulácia izotiokyanátových metabolitov v aktuálnom prsnom tkanive.
00:08:32 – Inhibícia kmeňových buniek rakoviny prsníka.
00:08:53 – Lekcia dejepisu: Brassica bola preukázaná ako zdraviu prospešné už v starovekom Ríme.
00:09:16 – Schopnosť sulforafanu zvyšovať vylučovanie karcinogénov (benzén, akroleín).
00:09:51 – NRF2 ako genetický prepínač prostredníctvom prvkov antioxidačnej odozvy.
00:10:10 – Ako aktivácia NRF2 zvyšuje vylučovanie karcinogénu cez glutatión-S-konjugáty.
00:10:34 – Ružičkový kel zvyšuje glutatión-S-transferázu a znižuje poškodenie DNA.
00:11:20 – Nápoj z brokolicových klíčkov zvyšuje vylučovanie benzénu o 61 %.
00:13:31 – Homogenát brokolicových klíčkov zvyšuje antioxidačné enzýmy v horných dýchacích cestách.
00:15:45 – Spotreba krížovej zeleniny a úmrtnosť na srdcové choroby.
00:16:55 – Prášok z brokolicových klíčkov zlepšuje krvné lipidy a celkové riziko srdcových chorôb u diabetikov 2. typu.
00:19:04 – Začiatok úseku starnutia.
00:19:21 – Diéta obohatená o sulforafan zvyšuje životnosť chrobákov z 15 na 30 % (za určitých podmienok).
00:20:34 – Význam nízkeho zápalu pre dlhovekosť.
00:22:05 – Zdá sa, že krížová zelenina a prášok z brokolicových klíčkov znižujú množstvo zápalových markerov u ľudí.
00:23:40 – Rekapitulácia v polovici videa: rakovina, starnutie
00:24:14 – Štúdie na myšiach naznačujú, že sulforafan môže zlepšiť adaptívnu imunitnú funkciu v starobe.
00:25:18 – Sulforaphane zlepšil rast vlasov u myšieho modelu plešatenia. Snímka o 00:26:10.
00:26:30 – Začiatok sekcie mozog a správanie.
00:27:18 – Vplyv extraktu z brokolicových klíčkov na autizmus.
00:27:48 – Účinok glukorafanínu na schizofréniu.
00:28:17 – Začiatok diskusie o depresii (pravdepodobný mechanizmus a štúdie).
00:31:21 – Štúdia na myšiach s použitím 10 rôznych modelov depresie vyvolanej stresom ukazuje, že sulforafan je podobne účinný ako fluoxetín (prozac).
00:32:00 – Štúdia ukazuje, že priame požitie glukorafanínu u myší je podobne účinné pri prevencii depresie zo sociálneho modelu stresu.
00:33:01 – Začiatok sekcie neurodegenerácie.
00:33:30 – Sulforafán a Alzheimerova choroba.
00:33:44 – Sulforaphane a Parkinsonova choroba.
00:33:51 - Sulforaphane a Hungtingtonova choroba.
00:34:13 – Sulforaphane zvyšuje proteíny tepelného šoku.
00:34:43 – Začiatok sekcie traumatického poranenia mozgu.
00:35:01 – Sulforaphane podaný ihneď po TBI zlepšuje pamäť (štúdia na myšiach).
00:35:55 – Sulforafán a plasticita neurónov.
00:36:32 – Sulforaphane zlepšuje učenie v modeli diabetu typu II u myší.
00:37:19 – Sulforaphane a Duchennova svalová dystrofia.
00:37:44 – Inhibícia myostatínu vo svalových satelitných bunkách (in vitro).
00:38:06 – Neskoré video rekapitulácia: úmrtnosť a rakovina, poškodenie DNA, oxidačný stres a zápal, vylučovanie benzénu, kardiovaskulárne ochorenia, diabetes typu II, účinky na mozog (depresia, autizmus, schizofrénia, neurodegenerácia), dráha NRF2.
00:40:27 – Myšlienky na to, ako zistiť dávku brokolicových klíčkov alebo sulforafanu.
00:41:01 – Anekdoty o klíčení doma.
00:43:14 – O teplotách varenia a aktivite sulforafanu.
00:43:45 – Premena sulforafanu z glukorafanínu črevnými baktériami.
00:44:24 – Doplnky fungujú lepšie v kombinácii s aktívnou myrozinázou zo zeleniny.
00:44:56 – Techniky varenia a krížová zelenina.
00:46:06 – Izotiokyanáty ako struma.
Nrf2 je transkripčný faktor, ktorý hrá dôležitú úlohu v bunkovom antioxidačnom obrannom systéme ľudského tela. Prvok reagujúci na antioxidant alebo ARE je regulačný mechanizmus génov. Mnohé výskumné štúdie preukázali, že Nrf2 alebo faktor 2 súvisiaci s NF-E2 reguluje širokú škálu génov riadených ARE v niekoľkých typoch buniek. Zistilo sa tiež, že Nrf2 hrá zásadnú úlohu pri bunkovej ochrane a antikarcinogenite, čo dokazuje, že Nrf2 môže byť účinnou liečbou pri liečbe neurodegeneratívnych ochorení a rakovín, o ktorých sa predpokladá, že sú spôsobené oxidačným stresom. Dr. Alex Jimenez DC, CCST Insight
Záverečné poznámky
Aj keď veľa otázok zostáva stále otvorených, dostupné experimentálne dôkazy jasne naznačujú, že Nrf2 je dôležitým hráčom pri udržiavaní mitochondriálnej homeostázy a štrukturálnej integrity. Táto úloha sa stáva obzvlášť kritickou v podmienkach oxidačného, elektrofilného a zápalového stresu, keď schopnosť upregulovať cytoprotektívne reakcie sprostredkované Nrf2 ovplyvňuje celkové zdravie a prežitie bunky a organizmu. Úloha Nrf2 v mitochondriálnej funkcii predstavuje ďalšiu vrstvu širokých cytoprotektívnych mechanizmov riadených týmto transkripčným faktorom. Pretože mnohé ľudské patologické stavy majú oxidačný stres, zápal a mitochondriálnu dysfunkciu ako základné zložky ich patogenézy, farmakologická aktivácia Nrf2 je prísľubom pre prevenciu a liečbu chorôb. Komplexné pochopenie presných mechanizmov, ktorými Nrf2 ovplyvňuje mitochondriálnu funkciu, je nevyhnutné pre racionálny návrh budúcich klinických štúdií a môže ponúknuť nové biomarkery na monitorovanie terapeutickej účinnosti.
Účelom vyššie uvedeného článku bolo diskutovať, ako aj demonštrovať vznikajúcu úlohu Nrf2 v mitochondriálnej funkcii. Nrf2, príp jadrový faktor erytroidný 2-príbuzný faktorje novým regulátorom bunkovej rezistencie voči oxidantom, ktorý môže prispievať k oxidačnému stresu, ovplyvňuje bunkovú funkciu a vedie k rozvoju toxicity, chronických ochorení a dokonca aj rakoviny. Zatiaľ čo produkcia oxidantov v ľudskom tele môže slúžiť na rôzne účely, vrátane bunkového delenia, zápalu, imunitnej funkcie, autofágie a stresovej reakcie, je nevyhnutné kontrolovať ich nadprodukciu, aby sa predišlo zdravotným problémom. Rozsah našich informácií je obmedzený na chiropraktické a zdravotné problémy chrbtice. Ak chcete prediskutovať túto tému, neváhajte sa opýtať Dr. Jimeneza alebo nás kontaktujte na adrese�915-850-0900 .
Bolesti chrbta�je jednou z najčastejších príčin invalidity a vymeškaných dní v práci na celom svete. Bolesti chrbta sú druhým najčastejším dôvodom návštevy lekára, prevyšujú ho iba infekcie horných dýchacích ciest. Približne 80 percent populácie aspoň raz za život zažije bolesť chrbta. Chrbtica je komplexná štruktúra pozostávajúca z kostí, kĺbov, väzov a svalov, okrem iných mäkkých tkanív. Z tohto dôvodu môžu zranenia a/alebo zhoršené stavy, ako naprherné disky, môže nakoniec viesť k príznakom bolesti chrbta. Športové zranenia alebo zranenia pri automobilových nehodách sú často najčastejšou príčinou bolesti chrbta, niekedy však môžu mať aj tie najjednoduchšie pohyby bolestivé následky. Našťastie alternatívne možnosti liečby, ako je chiropraktická starostlivosť, môžu pomôcť zmierniť bolesť chrbta pomocou úprav chrbtice a manuálnych manipulácií, čo v konečnom dôsledku zlepšuje úľavu od bolesti. �
Nrf2 podporuje aktiváciu skupiny antioxidačných a detoxikačných enzýmov a génov, ktoré chránia ľudský organizmus pred účinkami zdravotných problémov spojených so zvýšenou úrovňou oxidačného stresu, ako je Alzheimerova choroba. Ukázalo sa, že rôzne prírodné látky aktivujú dráhu Nrf2, ktorá môže pomôcť zvládnuť symptómy neurodegeneratívnych ochorení. Účelom nižšie uvedeného článku je diskutovať o kľúčovej úlohe Nrf2 spôsobenej chronickým zápalom.
abstraktné
Zápal je najčastejšou črtou mnohých chronických ochorení a komplikácií, pričom zohráva rozhodujúcu úlohu v karcinogenéze. Niekoľko štúdií preukázalo, že Nrf2 prispieva k protizápalovému procesu organizovaním náboru zápalových buniek a reguláciou génovej expresie prostredníctvom prvku antioxidačnej odozvy (ARE). Signálna dráha Keap1 (Kelch-like ECH-asociated protein)/Nrf2 (NF-E2 p45-related factor 2)/ARE hlavne reguluje protizápalovú génovú expresiu a inhibuje progresiu zápalu. Preto sa identifikácia nových protizápalových fytochemikálií závislých od Nrf2 stala kľúčovým bodom pri objavovaní liekov. V tomto prehľade diskutujeme o členoch signálnej dráhy Keap1/Nrf2/ARE a jej downstream génoch, účinkoch tejto dráhy na zvieracie modely zápalových ochorení a o presluchoch s dráhou NF-kB. Okrem toho diskutujeme aj o regulácii zápalu NLRP3 pomocou Nrf2. Okrem toho sumarizujeme súčasný scenár vývoja protizápalových fytochemikálií a ďalších, ktoré sprostredkovávajú signálnu dráhu Nrf2/ARE.
Zápal je komplexný proces, ku ktorému dochádza, keď sú tkanivá infikované alebo poškodené škodlivými stimulmi, ako sú patogény, poškodenie alebo dráždivé látky. Na tejto ochrannej reakcii sa podieľajú imunitné bunky, krvné cievy a molekulárne mediátory [1]. Zápal je tiež patologický jav spojený s rôznymi chorobnými stavmi vyvolanými najmä fyzikálnymi, chemickými, biologickými a psychologickými faktormi. Cieľom zápalu je obmedziť a odstrániť príčiny bunkového poškodenia, vyčistiť a/alebo absorbovať nekrotické bunky a tkanivá a iniciovať opravu tkaniva. Rozlišujú sa dve odlišné formy zápalu: akútny a chronický. Akútny zápal je samoobmedzujúci a prospešný pre hostiteľa, ale dlhotrvajúci chronický zápal je bežnou črtou mnohých chronických ochorení a komplikácií. Priama infiltrácia mnohými mononukleárnymi imunitnými bunkami, ako sú monocyty, makrofágy, lymfocyty a plazmatické bunky, ako aj produkcia zápalových cytokínov vedú k chronickému zápalu. Uznáva sa, že chronický zápal hrá rozhodujúcu úlohu v karcinogenéze [2]. Vo všeobecnosti sa pri normálnom zápalovom procese vzájomne ovplyvňujú prozápalové aj protizápalové signálne dráhy.
Pri patologickom zápalovom procese sa najskôr aktivujú žírne bunky, monocyty, makrofágy, lymfocyty a iné imunitné bunky. Potom sa bunky privedú na miesto poškodenia, čo vedie k vytvoreniu reaktívnych foriem kyslíka (ROS), ktoré poškodzujú makromolekuly vrátane DNA. Súčasne tieto zápalové bunky tiež produkujú veľké množstvo zápalových mediátorov, ako sú cytokíny, chemokíny a prostaglandíny. Tieto mediátory ďalej získavajú makrofágy na lokalizované miesta zápalu a priamo aktivujú viaceré kaskády prenosu signálu a transkripčné faktory spojené so zápalom. Signálne dráhy NF-KB (jadrový faktor kappa B), MAPK (mitogénom aktivovaná proteínkináza) a JAK (janus kináza)-STAT (prevodníky signálu a aktivátory transkripcie) sa podieľajú na vývoji klasickej dráhy zápalu [3], [4], [5]. Predchádzajúce štúdie odhalili, že transkripčný faktor Nrf2 (NF-E2 faktor 45 súvisiaci s p2) reguluje expresiu detoxikačných enzýmov fázy II vrátane NADPH, NAD(P)H chinónoxidoreduktázy 1, glutatiónperoxidázy, feritínu, hemoxygenázy-1 (HO -1) a antioxidačné gény, ktoré svojimi protizápalovými účinkami chránia bunky pred rôznymi poraneniami, a tak ovplyvňujú priebeh ochorenia [6], [7], [8].
Vzhľadom na tieto pozoruhodné zistenia, vývoj cielených terapeutických liekov na zápalové ochorenia prostredníctvom signálnych dráh pritiahol v posledných rokoch veľký záujem. V tomto prehľade sumarizujeme výskum signálnej dráhy Keap1 (Kelch-like ECH related protein)/Nrf2 (NF-E2 p45-related factor 2)/ARE (antioxidačná odozva element) signálnej dráhy pri zápale.
Štruktúra a regulácia Nrf2
Regulácia Keap1-Dependent Nrf2
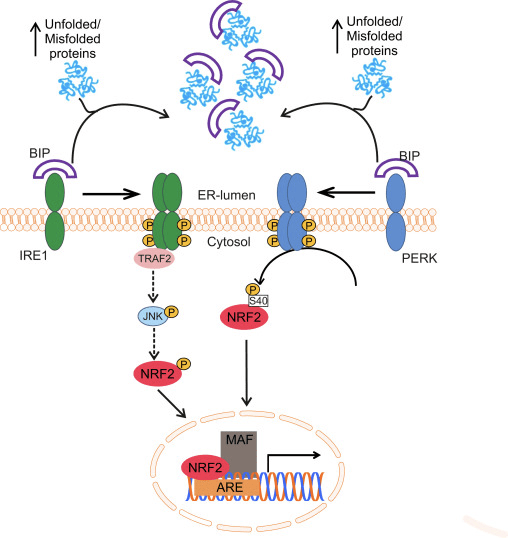
Nrf2 patrí do podrodiny Cap �n� Collar (CNC) a zahŕňa sedem funkčných domén Neh (homológia Nrf2-ECH) 1 až Neh7 [9], [10]. Neh1 je doména CNC-bZIP, ktorá umožňuje Nrf2 heterodimerizovať s malým muskuloaponeurotickým fibrosarkómovým (Maf) proteínom, DNA a ďalšími transkripčnými partnermi, ako aj vytvárať jadrový komplex s enzýmom konjugujúcim ubikvitín UbcM2 [11], [12]. Neh2 obsahuje dva dôležité motívy známe ako DLG a ETGE, ktoré sú nevyhnutné pre interakciu medzi Nrf2 a jeho negatívnym regulátorom Keap1 [13], [14].
Keap1 je substrátový adaptér pre E3 ubikvitín ligázu na báze cullinu, ktorý za normálnych podmienok inhibuje transkripčnú aktivitu Nrf2 prostredníctvom ubikvitinácie a proteazomálnej degradácie [15], [16], [17]. Domény KELCH homodiméru Keap1 sa viažu na motívy DLG a ETGE domény Nrf2-Neh2 v cytosóle, kde ETGE pôsobí ako pánt s vyššou afinitou a DLG pôsobí ako latch [18]. Pri oxidačnom strese alebo po vystavení aktivátorom Nrf2 sa Nrf2 disociuje z väzby Keap1 v dôsledku tiolovej modifikácie cysteínových zvyškov Keap1, čo v konečnom dôsledku zabraňuje ubikvitinácii Nrf2 a proteazomálnej degradácii [19]. Potom sa Nrf2 translokuje do jadra, heterodimerizuje sa s malými proteínmi Maf a transaktivuje ARE batériu génov (obr. 1A). Karboxy-koniec Neh3 pôsobí ako transaktivačná doména prostredníctvom interakcie s transkripčným koaktivátorom známym ako CHD6 (chromo-ATPáza/helikáza DNA viažuci proteín) [20]. Neh4 a Neh5 tiež pôsobia ako transaktivačné domény, ale viažu sa na iný transkripčný koaktivátor známy ako CBP (cAMP-response-element-binding protein-binding protein) [21]. Okrem toho Neh4 a Neh5 interagujú s jadrovým kofaktorom RAC3/AIB1/SRC-3, čo vedie k zvýšenej expresii génu ARE zacieleného na Nrf2 [22]. Neh5 má nukleárny exportný signál citlivý na redox, ktorý je rozhodujúci pre reguláciu a bunkovú lokalizáciu Nrf2 [23].
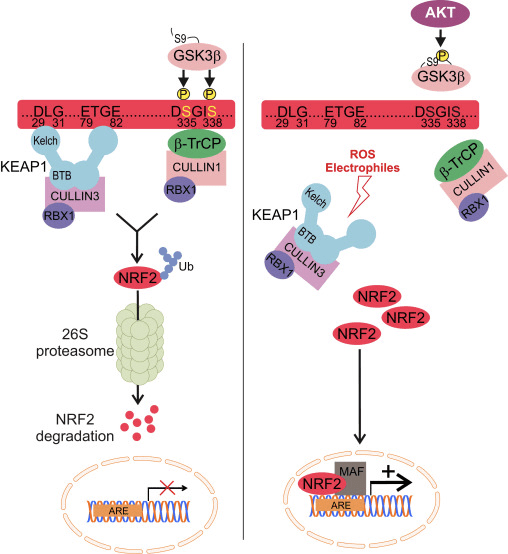
Obrázok 1 Keap1-dependentná a nezávislá regulácia Nrf2. (A) Za bazálnych podmienok je Nrf2 sekvestrovaný Keap1 jeho dvoma motívmi (ETGE a DLG), čo vedie k ubikvitinácii sprostredkovanej CUL3, po ktorej nasleduje degradácia proteazómu. Pri oxidačnom strese sa Nrf2 disociuje od Keap1, premiestňuje sa do jadra a aktivuje batériu génov ARE. (B) GSK3 fosforyluje Nrf2 a to uľahčuje rozpoznanie Nrf2 pomocou a-TrCP pre ubikvitináciu sprostredkovanú CUL1 a následnú degradáciu proteazómu. (C) p62 je sekvestrovaný Keap1, čo vedie k jeho autofagickej degradácii, uvoľneniu Nrf2 a zvýšenej signalizácii Nrf2.
Keap1-Nezávislá regulácia Nrf2
Nové dôkazy odhalili nový mechanizmus regulácie Nrf2, ktorý je nezávislý od Keap1. Neh6 doména Nrf2 bohatá na serín hrá kľúčovú úlohu v tejto regulácii tým, že sa viaže so svojimi dvoma motívmi (DSGIS a DSAPGS) na proteín obsahujúci repetíciu a-transducínu (a-TrCP) [24]. ?-TrCP je substrátový receptor pre komplex ubikvitín ligázy Skp1�Cul1�Rbx1/Roc1, ktorý sa zameriava na Nrf2 na ubikvitináciu a proteazomálnu degradáciu. Glykogénsyntáza kináza-3 je kľúčovým proteínom zapojeným do stabilizácie a regulácie Nrf1 nezávislej od Keap2; fosforyluje Nrf2 v doméne Neh6, aby sa uľahčilo rozpoznanie Nrf2 pomocou a-TrCP a následná degradácia proteínu [25] (obr. 1B).
Iné regulátory Nrf2
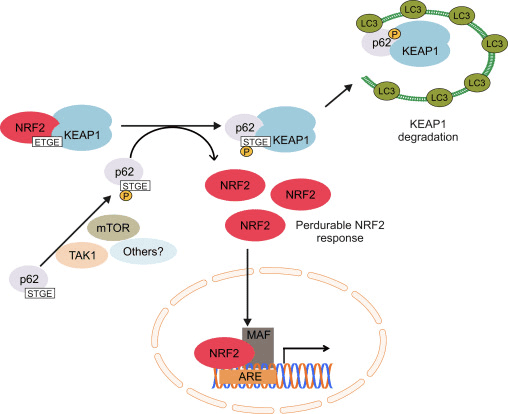
Ďalšia línia dôkazov odhalila nekanonickú dráhu aktivácie Nrf62 závislej od p2, v ktorej p62 sekvestruje Keap1 k autofagickej degradácii, ktorá nakoniec vedie k stabilizácii Nrf2 a transaktivácii génov závislých od Nrf2 [26], [27], [ 28], [29] (obr. 1C).
Hromadné dôkazy naznačujú, že niekoľko miRNA hrá dôležitú úlohu v regulácii aktivity Nrf2 [30]. Sangokoya a kol. [31] preukázali, že miR-144 priamo znižuje aktivitu Nrf2 v lymfoblastovej bunkovej línii K562, primárnych ľudských erytroidných progenitorových bunkách a retikulocytoch kosáčikovitej anémie. Ďalšia zaujímavá štúdia na ľudských epiteliálnych bunkách prsníka preukázala, že miR-28 inhibuje Nrf2 prostredníctvom mechanizmu nezávislého od Keap1 [32]. Podobne miRNA ako miR-153, miR-27a, miR-142-5p a miR144 downregulujú expresiu Nrf2 v neurónovej bunkovej línii SH-SY5Y [33]. Singh a kol. [34] preukázali, že ektopická expresia miR-93 znižuje expresiu génov regulovaných Nrf2 v modeli karcinogenézy prsníka u potkanov indukovanom 17p-estradiolom (E2).
Nedávny objav z nášho laboratória identifikoval endogénny inhibítor Nrf2 známy ako retinoický X receptor alfa (RXR?). RXR? je jadrový receptor, interaguje s doménou Neh7 Nrf2 (aminokyselinové zvyšky 209-316) prostredníctvom svojej DNA-väzbovej domény (DBD) a špecificky inhibuje aktivitu Nrf2 v jadre. Okrem toho sa uvádza, že endogénnymi inhibítormi aktivity Nrf2 sú aj iné jadrové receptory, ako je receptor-a aktivovaný peroxizómovým proliferátorom, ERp, receptor-a súvisiaci s estrogénom a glukokortikoidné receptory [9], [10].
Protizápalová úloha osi Nrf2/HO-1
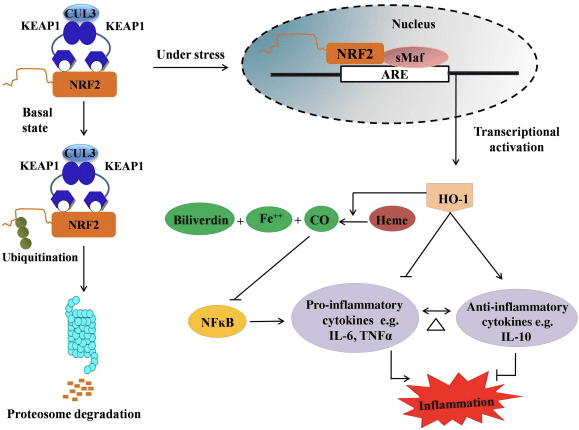
HO-1 je indukovateľná izoforma a enzým obmedzujúci rýchlosť, ktorý katalyzuje degradáciu hému na oxid uhoľnatý (CO) a voľné železo a biliverdín na bilirubín. Enzymatická degradácia prozápalového voľného hemu, ako aj produkcia protizápalových zlúčenín, ako je CO a bilirubín, zohrávajú hlavnú úlohu pri udržiavaní ochranných účinkov HO-1 (obr. 2).
Obrázok 2 Prehľad dráhy Nrf2/HO-1. Za bazálnych podmienok sa Nrf2 viaže na svoj represor Keap1, čo vedie k ubikvitinácii, po ktorej nasleduje degradácia proteazómu. Počas oxidačného stresu sa voľný Nrf2 translokuje do jadra, kde sa dimerizuje s členmi malej rodiny Maf a viaže sa na gény ARE, ako je HO-1. Upregulovaná HO-1 katalyzuje hem na CO, bilirubín a voľné železo. CO pôsobí ako inhibítor NF-kB dráhy, čo vedie k zníženej expresii prozápalových cytokínov, zatiaľ čo bilirubín pôsobí aj ako antioxidant. Okrem toho HO-1 priamo inhibuje prozápalové cytokíny, ako aj aktiváciu protizápalových cytokínov, čo vedie k vyrovnaniu zápalového procesu.
Nrf2 indukuje gén HO-1 zvýšením expresie mRNA a proteínu a je to jeden z klasických génov regulovaných Nrf2, ktorý sa široko používa v mnohých štúdiách in vitro a in vivo. Niekoľko štúdií preukázalo, že HO-1 a jeho metabolity majú významné protizápalové účinky sprostredkované Nrf2. Zvýšenie expresie HO-1, ktoré je sprostredkované aktivovaným Nrf2, vedie k inhibícii signalizácie NFkB, čo vedie k zníženiu poškodenia črevnej sliznice a dysfunkcii tesného spojenia v modeli transplantácie pečene samcov Sprague-Dawley potkanov [35]. Upregulácia expresie HO-2 závislej od Nrf1 môže chrániť myoblasty odvodené od myší C2C12 pred cytotoxicitou H2O2 [36]. HO-2 závislá od Nrf1 má vplyv na zápalové reakcie sprostredkované lipopolysacharidmi (LPS) v penových bunkových makrofágoch odvodených z RAW264.7 alebo myších peritoneálnych makrofágov. Aktivita Nrf2 znecitlivila fenotyp penových makrofágov a zabránila nemiernemu zápalu makrofágov, ktoré hrajú dôležitú úlohu v progresii aterosklerózy [37]. Os Nrf2/HO-1 ovplyvňuje LPS indukované myšie BV2 mikrogliálne bunky a myšie hipokampálne HT22 bunky s dopadom na neurozápal. Upregulácia expresie HO-1 prostredníctvom dráhy Nrf2 v myších mikrogliálnych bunkách BV2, ktoré bránia bunkovú smrť myších hipokampálnych buniek HT22 [38]. Okrem toho hybridné molekuly na báze kobaltu (HYCO), ktoré kombinujú induktor Nrf2 s uvoľňovačom oxidu uhoľnatého (CO), zvyšujú expresiu Nrf2/HO-1, uvoľňujú CO a prejavujú protizápalovú aktivitu in vitro. HYCO tiež upregulujú tkanivové HO-1 a dodávajú CO do krvi po podaní in vivo, čo podporuje ich potenciálne použitie proti zápalovým stavom [39]. Upregulácia Nrf2/HO-1 znižuje zápal zvýšením efferocytárnej aktivity myších makrofágov liečených taurínchloramínmi [40]. Celkovo vyššie vysvetlené experimentálne modely odhalili, že os Nrf2 / HO-1 hrá hlavnú úlohu v protizápalovej funkcii, čo naznačuje, že Nrf2 je terapeutickým cieľom pri ochoreniach spojených so zápalom.
Okrem toho vedľajšie produkty HO-1, ako je CO, bilirubín, pôsobí ako silný antioxidant počas oxidačného stresu a poškodenia buniek [41], [42]; potláča autoimunitnú encefalomyelitídu a hepatitídu [43], [44]; a chráni myši a potkany pred endotoxickým šokom tým, že bráni tvorbe iNOS a NO [45], [46], [47]. Bilirubín navyše znižuje aktiváciu a dysfunkciu endotelu [48]. Je zaujímavé, že bilirubín znižuje transmigráciu endotelových leukocytov cez adhéznu molekulu-1 [49]. Tieto špecifické odkazy naznačujú, že nielen HO-1 pôsobí ako silné protizápalové činidlo, ale aj jeho metabolity.
Zápalové mediátory a enzýmy inhibované Nrf2
Cytokíny a chemokíny
Cytokíny sú proteíny a polypeptidy s nízkou molekulovou hmotnosťou vylučované rôznymi bunkami; regulujú rast buniek, diferenciáciu a imunitnú funkciu a podieľajú sa na zápaloch a hojení rán. Cytokíny zahŕňajú interleukíny (IL), interferóny, tumor nekrotizujúci faktor (TNF), faktor stimulujúci kolónie, chemokíny a rastové faktory. Niektoré cytokíny sa považujú za prozápalové mediátory, zatiaľ čo iné majú protizápalové funkcie. Vystavenie oxidačnému stresu vedie k nadprodukcii cytokínov, čo spôsobuje oxidačný stres v cieľových bunkách. Keď je NF-kB aktivovaný oxidačným stresom, je nadprodukovaných niekoľko prozápalových cytokínov. Okrem toho prozápalový oxidačný stres spôsobuje ďalšiu aktiváciu NF-kB a nadprodukciu cytokínov. Dôležitú úlohu pri narušení tohto cyklu zohráva aktivácia systému Nrf2/ARE. Chemokíny sú skupinou malých cytokínov, ktorých hlavnou úlohou je riadiť migráciu zápalových buniek. Fungujú hlavne ako chemoatraktanty pre leukocyty, monocyty, neutrofily a iné efektorové bunky.
Uvádza sa, že aktivácia Nrf2 zabraňuje LPS-indukovanej transkripčnej upregulácii prozápalových cytokínov, vrátane IL-6 a IL-1a. [50]. IL-1? a produkcia IL-6 je tiež zvýšená v Nrf2?/? myši s kolitídou vyvolanou dextránsulfátom [51], [52]. Nrf2 inhibuje produkciu downstream IL-17 a iných zápalových faktorov Th1 a Th17 a potláča chorobný proces v experimentálnom modeli roztrúsenej sklerózy, autoimunitnej encefalitídy [53]. Antioxidačné gény HO-2, NQO-1, Gclc a Gclm závislé od Nrf1 blokujú TNF-a, IL-6, monocytový chemoatraktantový proteín-1 (MCP1), zápalový proteín makrofágov-2 (MIP2) a zápalové mediátorov. Ale v prípade Nrf2-knockout myší sa protizápalový účinok neprejavuje [54]. Peritoneálne neutrofily z myší s knockoutom Nrf2 liečených LPS majú významne vyššie hladiny cytokínov (TNF-a a IL-6) a chemokínov (MCP1 a MIP2) ako bunky divokého typu (WT) [54]. In vitro prenos génu Nrf2 do buniek hladkého svalstva ľudskej a králičej aorty potláča sekréciu MCP1 [8], [55] a expresia HO-2 závislá od Nrf1 potláča NF-?B a MCP-1 stimulované TNF-a sekrécie v endotelových bunkách ľudskej pupočníkovej žily [56]. Tieto zistenia naznačujú, že v reakcii na zápalové stimuly upregulácia signalizácie Nrf2 inhibuje nadprodukciu prozápalových cytokínov a chemokínov, ako aj obmedzuje aktiváciu NF-kB.
Molekuly bunkovej adhézie
Bunkové adhézne molekuly (CAM) sú proteíny, ktoré sa viažu s bunkami alebo s extracelulárnou matricou. Nachádzajú sa na bunkovom povrchu a podieľajú sa na rozpoznávaní buniek, aktivácii buniek, prenose signálu, proliferácii a diferenciácii. Spomedzi CAM sú ICAM-1 a VCAM-1 dôležitými členmi imunoglobulínovej superrodiny. ICAM-1 je prítomný v nízkych koncentráciách v membránach leukocytov a endotelových buniek. Po stimulácii cytokínom sa koncentrácia výrazne zvyšuje. ICAM-1 môže byť indukovaný IL-1 a TNF a je exprimovaný vaskulárnym endotelom, makrofágmi a lymfocytmi. Je to ligand pre integrín, receptor nachádzajúci sa na leukocytoch. Keď je aktivovaný ICAM-1-integrínový mostík, leukocyty sa viažu na endotelové bunky a potom migrujú do subendotelových tkanív [57]. VCAM-1 sprostredkováva adhéziu lymfocytov, monocytov, eozinofilov a bazofilov k vaskulárnemu endotelu a prispieva k získavaniu leukocytov, čo v konečnom dôsledku vedie k poškodeniu tkaniva v dôsledku oxidačného stresu. Nrf2 inhibuje promotorovú aktivitu VCAM-1 [58]. Nrf2-regulovaný downstream gén HO-1 môže ovplyvniť expresiu E-selektínu a VCAM-1, adhéznych molekúl asociovaných s endotelovými bunkami [59]. Pľúcna expresia niekoľkých CAM, ako napríklad CD-14, TREM1, SELE, SELP a VCAM-1, je výrazne vyššia v Nrf2a/? myšiach ako u myší Nrf2+/+ [60]. Nrf2 v ľudských endotelových bunkách aorty potláča expresiu VCAM-1 indukovanú TNF-a a interferuje s adhéziou monocytových buniek U937 indukovanou TNF-a [8]. Nadmerná expresia Nrf2 tiež inhibuje expresiu génu VCAM-1 indukovanú TNF-a v ľudských mikrovaskulárnych endotelových bunkách [61]. Zistilo sa, že prirodzene sa vyskytujúci antioxidant kyselina 3-hydroxyantranilová (HA), jeden z metabolitov l-tryptofánu tvorený in vivo metabolickou cestou známou ako kynurenínová dráha počas zápalu alebo infekcie, indukuje expresiu HO-1 a stimuluje Nrf2 v ľudskom pupočníkovom systéme. žilové endotelové bunky (HUVEC). Expresia HO-2 závislá od Nrf1 indukovaná HA inhibuje sekréciu MCP-1, expresiu VCAM-1 a aktiváciu NF-kB spojenú s vaskulárnym poškodením a zápalom pri ateroskleróze [56]. Antiproliferatívny a protizápalový syntetický chalkónový derivát 2a,4a,6a-tris(metoxymetoxy)chalkón inhibuje ICAM-1, prozápalový cytokín IL-1a a TNF-a. expresia v tkanive hrubého čreva od myší liečených kyselinou trinitrobenzénsulfónovou [62]. Upregulácia Nrf2 inhibuje TNF-a-indukovanú expresiu ICAM-1 v ľudských retinálnych pigmentových epiteliálnych bunkách ošetrených lykopénom [63]. Všetky tieto štúdie naznačujú, že Nrf2 hrá kľúčovú úlohu v zápalovom procese reguláciou migrácie a infiltrácie zápalových buniek do zapáleného tkaniva.
Matrixové metaloproteinázy (MMP)
MMP sú široko prítomné v extracelulárnej matrici a podieľajú sa na fyziologických a patologických procesoch, ako je bunková proliferácia, migrácia, diferenciácia, hojenie rán, angiogenéza, apoptóza a metastázy nádorov. Bolo publikované, že os Nrf2/HO-1 inhibuje MMP-9 v makrofágoch a MMP-7 v ľudských črevných epiteliálnych bunkách, čo je prospešné pri liečbe zápalového ochorenia čriev [62], [64]. Poškodenie kože spôsobené UV žiarením je závažnejšie pri knockoutovaní Nrf2 ako u myší WT a hladina MMP-9 je výrazne vyššia, čo naznačuje, že Nrf2 znižuje expresiu MMP-9. Preto sa Nrf2 považuje za ochranný prostriedok proti UV žiareniu [65]. Ďalšia štúdia tiež uvádza, že downregulovaná transkripčná aktivácia MMP-9 pri invázii nádorových buniek a zápale je regulovaná prostredníctvom inhibície NF-kB signálnej dráhy [66]. Pri traumatickom poškodení miechy sa signálna dráha NF-kB tiež podieľa na regulácii hladín mRNA MMP-9 [67]. Preto je pri zápale regulácia MMP ovplyvnená priamo dráhou Nrf2 alebo nepriamo prostredníctvom dráhy NF-kB ovplyvnenej Nrf2.
Cyklooxygenáza-2 (COX2) a indukovateľná syntáza oxidu dusnatého (INOS)
Séria experimentov na myšiach s knockoutom Nrf2 preukázala jeho kľúčovú úlohu pri zápale a regulácii prozápalových génov, ako sú COX-2 a iNOS. Prvýkrát Khor a spol. hlásili zvýšenú expresiu prozápalových cytokínov, ako sú COX-2 a iNOS v tkanivách hrubého čreva Nrf2a/? myši v porovnaní s myšami WT Nrf2+/+, čo naznačuje, že Nrf2 potláča ich aktivitu [51]. Ďalšia správa o predbežnej úprave sulforafanom, jedným z dobre známych aktivátorov Nrf2 prítomných v krížovej zelenine, preukázala svoj protizápalový účinok inhibície expresie TNF-a, IL-1a, COX-2 a iNOS na oboch mRNA a hladiny proteínov v primárnych peritoneálnych makrofágoch z myší Nrf2+/+ v porovnaní s hladinami z Nrf2a/? myši [68]. Podobne hipokampus Nrf2-knockout myší so zápalom indukovaným LPS tiež vykazuje vyššiu expresiu zápalových markerov, ako sú iNOS, IL-6 a TNF-? než WT myši [69]. Podobne myši s knockoutom Nrf2 sú precitlivené na oxidačný stres vyvolaný 1-metyl-4-fenyl-1,2,3,6-tetrahydropyridínom, ako aj vykazujú zvýšené hladiny mRNA a proteínov zápalových markerov, ako sú COX-2, iNOS IL-6 a TNF-a [70]. Navyše pečeň z Nrf2?/? myši vystavené diéte s deficitom metionínu a cholínu majú ~ 5-krát vyššiu expresiu mRNA Cox2 a iNOS ako myši od WT myší na rovnakej diéte, čo naznačuje protizápalovú úlohu Nrf2 [71]. Nedávno Kim a spol. preukázali, že fytochemický etylpyruvát uplatňuje svoje protizápalové a antioxidačné účinky znížením expresie iNOS prostredníctvom signalizácie Nrf2 v bunkách BV2. Ukázali, že etylpyruvát indukuje jadrovú translokáciu Nrf2, čo v konečnom dôsledku inhibuje interakciu medzi p65 a p300, čo vedie k zníženej expresii iNOS [72]. Okrem toho karbazolový analóg LCY-2-CHO aktivuje Nrf2 a spôsobuje jeho jadrovú translokáciu, čo vedie k potlačeniu expresie COX2 a iNOS [73] v bunkách hladkého svalstva aorty potkana.
Paradoxná úloha Nrf2 v regulácii aktivity iIflammasómu NLRP3
Rodina NLR, pyrínová doména obsahujúca 3 (NLRP3) zápalové bunky, je multiproteínový komplex, ktorý funguje ako receptor na rozpoznávanie patogénov (PRR) a rozpoznáva široké spektrum signálov mikrobiálneho, oxidačného stresu, ako sú molekulárne vzory spojené s patogénmi (PAMP), poškodenie- asociované molekuly molekulového vzoru (DAMP) a ROS [74]. Aktivovaný zápal NLRP3 sprostredkuje štiepenie kaspázy-1 a sekréciu prozápalového cytokínu interleukínu-1? (IL-1?), ktorý v konečnom dôsledku indukuje proces bunkovej smrti známy ako pyroptóza, ktorý chráni hostiteľa pred širokým spektrom patogénov [75]. Aberantná aktivácia zápalu je však spojená s chorobami nesprávneho poskladania proteínov, ako sú prenosné spongiformné encefalopatie, Alzheimerova choroba, Parkinsonova choroba a tiež diabetes 2. typu [76], rakovina [77], dna a ateroskleróza [78].
Nedávne pozorovanie od skupiny Rong Hu o asociácii Nrf2 s negatívnou reguláciou zápalu zápalu odhalilo, že Nrf2 indukuje expresiu NQ1, ktorá vedie k inhibícii aktivácie zápalu NLRP3, štiepeniu kaspázy-1 a IL-1? generácie v makrofágoch. Okrem toho dobre známy aktivátor Nrf2, terc-butylhydrochinón (tBHQ), negatívne reguloval transkripciu NLRP3 aktiváciou ARE spôsobom závislým od Nrf2 [79]. Okrem vyššie uvedeného pozorovania sa tej istej skupine tiež zistilo, že dimetylfumarát (DMF) zabraňuje kolitíde indukovanej DSS prostredníctvom aktivácie signálnej dráhy Nrf2, ktorá sa podieľa na jadrovej translokácii Nrf2 a inhibícii zostavy zápalu NLRP3 [80].
Séria experimentov s použitím prírodných a syntetických zlúčenín tiež odhalila inhibičný účinok Nrf2 na aktiváciu zápalu NLRP3. Napríklad liečba epigalokatechín-3-galátom (EGCG) u myší s lupusovou nefritídou preukázala zníženie aktivácie obličkového zápalu NLRP3, ktorá je sprostredkovaná signálnou dráhou Nrf2 [81]. Podobne citral (3,7-dimetyl-2,6-oktadienal), hlavná účinná látka v čínskom bylinnom lieku Litsea cubeba, inhibuje aktiváciu zápalu NLRP3 prostredníctvom antioxidačnej signálnej dráhy Nrf2 v myšom modeli so zrýchleným a ťažkým lupusovým nefritídom (ASLN). [82]. Podobne biochanín chránil pred poškodením pečene vyvolaným LPS/GalN aktiváciou dráhy Nrf2 a inhibíciou aktivácie zápalu NLRP3 u samcov myší BALB/c [83]. Ďalej sa tiež ukázalo, že mangiferín up-reguluje expresiu Nrf2 a HO-1 spôsobom závislým od dávky a inhibuje LPS/D-GalN-indukovanú pečeňovú NLRP3, ASC, kaspázu-1, IL-1? a TNF-? výraz [84].
Napriek negatívnej regulácii NLRP3 pomocou Nrf2 tiež aktivuje funkciu zápalu NLRP3 a AIM2. Haitao Wen a kolegovia zistili, že Nrf2 ?/? myšie makrofágy preukázali defektnú aktiváciu zápalu NLRP3 a AIM2, ale nie zápalu NLRC4 [85]. Je zaujímavé, že toto pozorovanie zobrazuje neznáme funkcie Nrf2 v kontexte chorôb spojených so zápalom; preto je veľmi dôležité ďalej študovať, aby sa odhalil mechanizmus, v ktorom Nrf2 aktivuje inflammasómovú funkciu predtým, ako sa to považuje za terapeutický cieľ.
Potlačenie transkripcie prozápalových cytokínov pomocou Nrf2
Veľmi nedávny výskum založený na chromatínovej imunoprecipitácii (ChIP)-seq a ChIP-qPCR výsledky v myších makrofágoch odhalili, že Nrf2 sa viaže na promótorové oblasti prozápalových cytokínov, ako sú IL-6 a IL-1? a inhibuje získavanie RNA Pol II. Výsledkom je, že RNA Pol II nie je schopná spracovať transkripčnú aktiváciu IL-6 a IL-1a. čo v konečnom dôsledku vedie k inhibícii génovej expresie. Skupina Masayuki Yamamoto po prvýkrát odhalila nový mechanizmus, ktorým Nrf2 nielen transaktivuje svoje downstream gény prostredníctvom ARE, ale tiež potláča transkripčnú aktiváciu špecifických génov s ARE alebo bez neho prostredníctvom inhibície náboru RNA Pol II [50].
Presluchy medzi cestami Nrf2 a NF-KB
NF-kB je proteínový komplex zodpovedný za transkripciu DNA, ktorý sa nachádza takmer vo všetkých typoch živočíšnych buniek a podieľa sa na rôznych procesoch, ako je zápal, apoptóza, imunitná odpoveď, bunkový rast a vývoj. p65, proteín Rel z rodiny NF-kB, má transaktivačnú doménu, zatiaľ čo p50 ju nemá a na aktiváciu transkripcie vyžaduje heterodimerizáciu s proteínom Rel. Počas oxidačného stresu sa IkB kináza (IKK) aktivuje a spôsobuje fosforyláciu IkB, čo vedie k uvoľneniu a jadrovej translokácii NF-kB. NF-KB spôsobuje transkripciu prozápalových mediátorov, ako je IL-6, TNF-a, iNOS, IL-1 a intracelulárna adhézia COX-2.
Abnormálna regulácia NF-kB je spojená s reumatoidnou artritídou, astmou, zápalovým ochorením čriev a gastritídou vyvolanou infekciou Helicobacter pylori [86]. V súčasnosti sa predpokladá, že aktivita NF-kB ovplyvňuje signálnu dráhu Keapl/Nrf2/ARE hlavne v troch aspektoch: po prvé, Keap1 degraduje IKK? prostredníctvom ubikvitinácie, čím sa inhibuje aktivita NF-KB [87]. Po druhé, zápalový proces indukuje zápalové mediátory ako COX2 odvodené z cyklopentenón prostaglandínu 15d-PGJ2, silného elektrofilu, ktorý reaguje s Keap1 a aktivuje Nrf2, čím sa iniciuje transkripcia génu so súčasnou inhibíciou aktivity NF-kB [58], [88] ( Obr. 3 A, B). Po tretie, NF-kB sa môže kombinovať s kompetitívnym Nrf2 transkripčným koaktivátorom CBP [89], [90] (obr. 3 C, D).
Obrázok 3 Presluchy medzi dráhami Nrf2 a NF-KB. (A) Keap1 nasmeruje IKK na ubikvitináciu a degradáciu proteazómu sprostredkovanú CUL3, čo nakoniec vedie k inhibícii fosforylácie NF-kB a tento mechanizmus funguje aj ako kompetitívna väzba Nrf2 a IKK s Keap1. (B) Oxidačný stres aktivuje IKK, ktorý fosforyluje NF-kB, čo vedie k jeho translokácii do jadra a aktivácii prozápalových cytokínov, ako je COX-2. Koncový produkt COX-2 známy ako 15d-PGJ2 pôsobí ako induktor Nrf2, ktorý v konečnom dôsledku vedie k potlačeniu oxidačného stresu. (C) Nrf2 sa viaže so svojím transkripčným kofaktorom CBP spolu s malým Maf a iným transkripčným aparátom, aby inicioval génovú expresiu riadenú ARE. (D) Keď sa NF-kB viaže s CBP kompetitívnym spôsobom, inhibuje väzbu CBP s Nrf2, čo vedie k inhibícii transaktivácie Nrf2.
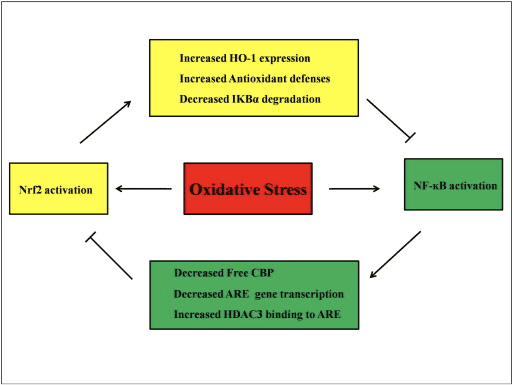
Predpokladá sa, že signálne dráhy Nrf2 a NF-kB interagujú pri kontrole transkripcie alebo funkcie downstream cieľových proteínov. Na odôvodnenie tohto predpokladu mnohé príklady ukazujú, že medzi členmi dráh Nrf2 a NF-kB dochádza k priamej alebo nepriamej aktivácii a inhibícii (obr. 4). V reakcii na LPS knockdown Nrf2 významne zvyšuje transkripčnú aktivitu NF-KB a transkripciu génu závislú od NF-KB, čo ukazuje, že Nrf2 bráni aktivite NF-KB [60], [91]. Okrem toho zvýšená expresia Nrf2-dependentného downstream HO-1 inhibuje aktivitu NF-kB. Keď sú bunky rakoviny prostaty krátko vystavené p-tochoferylsukcinátu, derivátu vitamínu E, expresia HO-1 je zvýšená. Koncové produkty HO-1 inhibujú jadrovú translokáciu NF-KB [92]. Tieto štúdie in vivo naznačujú, že Nrf2 negatívne reguluje signálnu dráhu NF-kB. LPS stimuluje NF-kB DNA väzbovú aktivitu a hladina p65 podjednotky NF-kB je výrazne vyššia v jadrových extraktoch z pľúc Nrf2a/? než od WT myší, čo naznačuje negatívnu úlohu Nrf2 pri aktivácii NF-kB. Navyše, Nrf2?/? fibroblasty myších embryí ošetrené LPS a TNF-? ukazujú výraznejšiu aktiváciu NF-KB spôsobenú aktiváciou IKK a I?B-? degradácia [60]. A klírens respiračného syncyciálneho vírusu je výrazne znížený, zatiaľ čo väzbová aktivita NF-KB DNA sa v Nrf2a/a zvyšuje. myši v porovnaní s myšami WT [93]. Pristane indukovaná lupusová nefritída u Nrf2?/? myši súbežne liečené sulforafanom majú vážne poškodenie obličiek a patologické zmeny, ako aj zvýšenú expresiu iNOS a aktiváciu NF-kB v porovnaní s WT, čo naznačuje, že Nrf2 zlepšuje lupusovú nefritídu inhibíciou signálnej dráhy NF-kB a odstránením ROS [94 ]. Aktivita NF-kB sa vyskytuje aj vtedy, keď sú bunky ošetrené induktorom Nrf2 spolu s LPS a TNF-a. Napríklad syntetický derivát chalkónu inhibuje aktiváciu NF-kB indukovanú TNF-a priamo aj nepriamo a čiastočne prostredníctvom indukcie expresie HO-1 v ľudských črevných epitelových bunkách HT-29 [62]. Potlačenie translokácie NF-kB a väzbovej aktivity DNA, ako aj potlačenie expresie iNOS v hepatocytoch sa zistilo, keď sa potkany F344 ošetrili 3H-1,2-ditiol-3-tiónom (D3T) [95]. Po spoločnej liečbe sulforafanom a LPS bola LPS-indukovaná expresia iNOS, COX-2 a TNF-? v Raw 264.7 makrofágoch je downregulovaný, čo naznačuje, že sulforafan má protizápalovú aktivitu prostredníctvom inhibície väzby NF-kB DNA [96]. Hoci bolo vykonaných niekoľko experimentálnych štúdií na vysvetlenie spojenia medzi dráhami Nrf2 a NF-kB, zostávajú protichodné výsledky. Medzi Nrf2 a NF-kB boli zaznamenané pozitívne aj negatívne regulácie [97]. Chemopreventívne elektrofily 3H-1,2-ditiol-3-tión, sulforafan a triterpenoid CDDO-Me zvyčajne aktivujú Nrf2 inhibíciou NF-kB a jeho downregulovaných génov [98], [99], [100]. Na rozdiel od toho sa ukázalo, že niekoľko látok alebo stavov, ako sú ROS, LPS, šmykové napätie toku, oxidovaný LDL a cigaretový dym, zvyšujú aktivitu Nrf2 aj NF-kB [97]. Okrem toho štúdie in vivo odhalili, že aktivita NF-kB je znížená v pečeni izolovanej z Nrf2a/p. myši a väzbová aktivita NF-KB je nižšia u Nrf2a/a. než u Nrf2+/+ myší [101]. Avšak ľudské aortálne endotelové bunky ošetrené adenovírusovým vektorom Nrf2 inhibujú NF-kB downstream gény bez ovplyvnenia aktivity NF-kB [8].
Obrázok 4 Regulačná slučka Nrf2 a NF-KB. Dráha Nrf2 inhibuje aktiváciu NF-kB tým, že bráni degradácii IkB-a. a zvýšenie expresie HO-1 a antioxidačnej obrany, ktorá neutralizuje ROS a detoxikačné chemikálie. V dôsledku toho je aktivácia NF-kB spojená s ROS potlačená. Podobne NF-KB-sprostredkovaná transkripcia znižuje aktiváciu Nrf2 redukciou�SÚgénová transkripcia a voľný proteín viažuci CREB súťažou s Nrf2 o CBP. Okrem toho NF-kB zvyšuje nábor histón deacetylázy (HDAC3) do oblasti ARE, a preto sa zabráni aktivácii transkripcie Nrf2.
Aktivácia signálnej dráhy Nrf2 hrá hlavnú úlohu pri expresii enzýmov a génov zapojených do detoxikácie reaktívnych oxidantov zvýšením antioxidačnej kapacity buniek v ľudskom tele. Aj keď je dnes k dispozícii veľa výskumných štúdií, regulačné mechanizmy v aktivácii Nrf2 nie sú úplne pochopené. Zistila sa aj možná úloha signálnej dráhy Nrf2 pri liečbe zápalu. Dr. Alex Jimenez DC, CCST Insight
Úloha Nrf2 pri zápalových ochoreniach
Štúdie in vivo ukázali, že Nrf2 hrá dôležitú úlohu pri zápalových ochoreniach postihujúcich rôzne systémy; tieto zahŕňajú gastritídu, kolitídu, artritídu, pneumóniu, poškodenie pečene, kardiovaskulárne ochorenie, neurodegeneratívne ochorenie a poškodenie mozgu. V týchto štúdiách Nrf2?/? zvieratá vykazovali závažnejšie príznaky zápalu a poškodenia tkaniva ako zvieratá WT. Preto sa predpokladá, že signálna dráha Nrf2 má ochranný účinok pri zápalových ochoreniach. Intratracheálna inštalácia prasacej pankreatickej elastázy indukuje chronickú obštrukčnú chorobu pľúc, najmä emfyzém. Myši s deficitom Nrf2 sú vysoko citlivé na emfyzém a znížená expresia HO-1, PrxI a antiproteázového génu SLPI sa vyskytuje v alveolárnych makrofágoch. Nrf2 sa považuje za kľúčový regulátor v makrofágmi sprostredkovanom obrannom systéme proti poraneniu pľúc [102]. Myši s deficitom Nrf2 s emfyzémom vyvolaným vystavením tabakovému dymu počas 6 mesiacov vykazujú zvýšený bronchoalveolárny zápal, upregulovanú expresiu markerov oxidačného stresu v alveolách a zvýšenú apoptózu alveolárnych septálnych buniek, čo naznačuje, že Nrf2 pôsobí proti emfyzému vyvolanému tabakom prostredníctvom zvýšenej expresie antioxidantu gény [102], [103]. Pri prerušení Nrf2 alergénom sprostredkovaný zápal dýchacích ciest a astma s použitím komplexu ovalbumínu vykazujú zvýšený zápal dýchacích ciest, hyperreaktivitu dýchacích ciest, hyperpláziu pohárikovitých buniek a vysoké hladiny Th2 v bronchoalveolárnom výplachu a splenocytoch, zatiaľ čo signálna dráha sprostredkovaná Nrf2 obmedzuje eozinofíliu dýchacích ciest hypersekrécia hlienu a hyperreaktivita dýchacích ciest, ako aj indukcia mnohých antioxidačných génov, ktoré bránia rozvoju astmy [104]. Injekcia karagénanu do pleurálnej dutiny vyvoláva zápal pohrudnice a akumulácia 15d-PGJ2 v zápalových bunkách Nrf2 je obmedzená na myšie peritoneálne makrofágy. Počas skorej fázy zápalu 15d-PGJ2 aktivuje Nrf2 a reguluje zápalový proces prostredníctvom indukcie HO-1 a PrxI. Štúdia tiež naznačila, že COX-2 má v ranej fáze protizápalový účinok produkciou 15d-PGJ2 [105]. Perorálne podávanie 1% dextránsulfátu sodného počas 1 týždňa indukuje kolitídu spojenú s histologickými zmenami, ktoré zahŕňajú skrátenie krýpt a infiltráciu zápalových buniek v tkanive hrubého čreva. Na ochranu črevnej integrity pri kolitíde by Nrf2 mohol hrať dôležitú úlohu reguláciou prozápalových cytokínov a indukciou detoxikačných enzýmov fázy II [51]. V myšom modeli LPS-indukovanej pľúcnej sepsy s knockoutom Nrf2 reguluje aktivita NF-kB vplyv zápalových cytokínov, ako sú COX-2, IL-113, IL-6 a TNFa. ktoré sú nevyhnutné na spustenie a podporu zápalu [60]. Nrf2 znižuje zápalové poškodenie reguláciou týchto zápalových faktorov. V týchto modeloch akútneho zápalu zvýšená regulácia antioxidačných enzýmov, prozápalových cytokínov a mediátorov signálnou dráhou Nrf2 znižuje zápalové poškodenie u zvierat WT. Je zaujímavé, že to bolo hlásené aj u myší s knockoutom Nrf2, u ktorých sú symptómy výrazne zhoršené v porovnaní s myšami WT.
Výskum protizápalových liekov závislých od Nrf2
V súhrne sme diskutovali o experimentoch, ktoré ukazujú, že signálna dráha Nrf2 hrá regulačnú úlohu v mnohých oblastiach zápalu, takže protizápalové látky závislé od Nrf2 sú dôležité na liečbu zápalových ochorení.
Rastliny boli mimoriadne bohatým zdrojom zlúčenín, ktoré aktivujú transkripčný faktor Nrf2, čo vedie k up-regulácii cytoprotektívnych génov. Nedávno sa uskutočnilo niekoľko štúdií na skúmanie účinkov rôznych protizápalových látok, väčšinou rastlinného pôvodu. Napríklad kurkumín je aktívnou zložkou kurkumy a v malom množstve sa nachádza aj v zázvore; izotiokyanáty, konkrétne fenylizotiokyanáty, sú z brokolice, zeleru a inej zeleniny; a antokyány sú z bobúľ a hrozna [124]. Štúdie ukázali, že všetky tieto látky sú nielen dobrými antioxidantmi, ale majú aj silné protizápalové účinky prostredníctvom indukcie Nrf2 [125], [126]. Preto vývoj nových protizápalových aktivátorov Nrf2 z rastlinného extraktu pritiahol veľký záujem v lekárskom výskume.
V posledných rokoch sa uskutočnilo mnoho pokusov na zvieratách na potvrdenie účinkov týchto zlúčenín. Artesunate sa používa hlavne pri ťažkej malárii, cerebrálnej malárii a reumatických autoimunitných ochoreniach; je účinný aj pri septickom poranení pľúc. Artesunát aktivuje expresiu Nrf2 a HO-1, ktorá znižuje prítok prozápalových cytokínov a leukocytov do tkaniva, aby sa zabránilo zápalu [127]. Predpokladá sa, že izovitexín, extrahovaný zo šupiek ryže Oryza sativa, má protizápalové a antioxidačné vlastnosti; hrá ochrannú úlohu proti akútnemu poškodeniu pľúc vyvolanému LPS aktiváciou dráhy Nrf2/HO-1 a inhibíciou MAPK a NF-?B [128]. Fimasartan, novo populárny blokátor receptora angiotenzínu II pôsobiaci na systém renín-angiotenzín, znižuje krvný tlak; použitie fimasartanu na liečbu myší s chirurgicky indukovanou unilaterálnou ureterálnou obštrukciou znižuje oxidačný stres, zápal a fibrózu prostredníctvom upregulácie Nrf2 a antioxidačnej dráhy a inhibíciou RAS a MAPK [129]. Sappanon je široko distribuovaný v juhovýchodnej Ázii, kde sa používa ako protichrípkový, antialergický a neuroprotektívny liek; aktivuje Nrf2 a inhibuje NF-kB, a tak môže byť prospešný pri liečbe chorôb súvisiacich s Nrf2 a/alebo NF-kB [130]. Bixin extrahovaný zo semien Bixin orellana sa používa na infekčné a zápalové ochorenia v Mexiku a Južnej Amerike; znižuje zápalové mediátory, presakovanie alveolárnych kapilár a oxidačné poškodenie spôsobom závislým od Nrf2, aby sa zmiernilo poškodenie pľúc vyvolané ventiláciou a obnovila sa normálna morfológia pľúc [131]. Iné rastlinné zlúčeniny, ako je epigalokatechín galát, sulforafan, resveratrol, lykopén a extrakt zo zeleného čaju, majú terapeutické účinky na zápalové ochorenia prostredníctvom signálnej dráhy Nrf2 [132], [133], [134]. Nedávno sa uvádza, že ďalšia fytochemikália, eriodictyol, ktorý je prítomný v citrusových plodoch, má protizápalové a antioxidačné účinky na poškodenie obličiek vyvolané cisplatinou a akútne poškodenie pľúc vyvolané sepsou reguláciou Nrf2, inhibíciou NF-KB a inhibíciou expresia cytokínov v makrofágoch [135], [136]. Mnohé fytochemikálie sú však veľmi sľubné pre prevenciu a liečbu rôznych ľudských chorôb a niektoré už vstúpili do štádia klinických skúšok (tabuľka 2).
Tieto rastlinné zlúčeniny aktivujú signálnu dráhu Nrf2 hlavne vo forme elektrofilných materiálov, ktoré modifikujú cysteínové zvyšky Keap1, čo vedie k voľnej väzbe jadrového Nrf2 s ARE, čo vedie k aktivácii transkripcie zodpovedajúceho génu.
Sulforaphane a jeho účinky na rakovinu, úmrtnosť, starnutie, mozog a správanie, srdcové choroby a ďalšie
Izotiokyanáty sú niektoré z najdôležitejších rastlinných zlúčenín, ktoré môžete získať vo svojej strave. V tomto videu pre nich robím najkomplexnejší prípad, aký kedy bol vyrobený. Krátka doba pozornosti? Preskočte na svoju obľúbenú tému kliknutím na jeden z časových bodov nižšie. Úplná časová os nižšie.
Kľúčové sekcie:
00:01:14 – Rakovina a úmrtnosť
00:19:04 – Starnutie
00:26:30 – Mozog a správanie
00:38:06 – Záverečná rekapitulácia
00:40:27 – Dávka
Úplná časová os:
00:00:34 – Predstavenie sulforafanu, hlavné zameranie videa.
00:01:14 – Spotreba krížovej zeleniny a zníženie úmrtnosti zo všetkých príčin.
00:02:12 – Riziko rakoviny prostaty.
00:02:23 – Riziko rakoviny močového mechúra.
00:02:34 – Riziko rakoviny pľúc u fajčiarov.
00:02:48 – Riziko rakoviny prsníka.
00:03:13 – Hypotetické: čo ak už máte rakovinu? (intervenčné)
00:03:35 – Pravdepodobný mechanizmus, ktorý riadi asociatívne údaje o rakovine a úmrtnosti.
00:04:38 – Sulforafán a rakovina.
00:05:32 – Dôkazy na zvieratách ukazujúce silný účinok extraktu z výhonkov brokolice na vývoj nádoru močového mechúra u potkanov.
00:06:06 – Účinok priamej suplementácie sulforafanu u pacientov s rakovinou prostaty.
00:07:09 – Bioakumulácia izotiokyanátových metabolitov v aktuálnom prsnom tkanive.
00:08:32 – Inhibícia kmeňových buniek rakoviny prsníka.
00:08:53 – Lekcia dejepisu: Brassica bola preukázaná ako zdraviu prospešné už v starovekom Ríme.
00:09:16 – Schopnosť sulforafanu zvyšovať vylučovanie karcinogénov (benzén, akroleín).
00:09:51 – NRF2 ako genetický prepínač prostredníctvom prvkov antioxidačnej odozvy.
00:10:10 – Ako aktivácia NRF2 zvyšuje vylučovanie karcinogénu cez glutatión-S-konjugáty.
00:10:34 – Ružičkový kel zvyšuje glutatión-S-transferázu a znižuje poškodenie DNA.
00:11:20 – Nápoj z brokolicových klíčkov zvyšuje vylučovanie benzénu o 61 %.
00:13:31 – Homogenát brokolicových klíčkov zvyšuje antioxidačné enzýmy v horných dýchacích cestách.
00:15:45 – Spotreba krížovej zeleniny a úmrtnosť na srdcové choroby.
00:16:55 – Prášok z brokolicových klíčkov zlepšuje krvné lipidy a celkové riziko srdcových chorôb u diabetikov 2. typu.
00:19:04 – Začiatok úseku starnutia.
00:19:21 – Diéta obohatená o sulforafan zvyšuje životnosť chrobákov z 15 na 30 % (za určitých podmienok).
00:20:34 – Význam nízkeho zápalu pre dlhovekosť.
00:22:05 – Zdá sa, že krížová zelenina a prášok z brokolicových klíčkov znižujú množstvo zápalových markerov u ľudí.
00:23:40 – Rekapitulácia v polovici videa: rakovina, starnutie
00:24:14 – Štúdie na myšiach naznačujú, že sulforafan môže zlepšiť adaptívnu imunitnú funkciu v starobe.
00:25:18 – Sulforaphane zlepšil rast vlasov u myšieho modelu plešatenia. Snímka o 00:26:10.
00:26:30 – Začiatok sekcie mozog a správanie.
00:27:18 – Vplyv extraktu z brokolicových klíčkov na autizmus.
00:27:48 – Účinok glukorafanínu na schizofréniu.
00:28:17 – Začiatok diskusie o depresii (pravdepodobný mechanizmus a štúdie).
00:31:21 – Štúdia na myšiach s použitím 10 rôznych modelov depresie vyvolanej stresom ukazuje, že sulforafan je podobne účinný ako fluoxetín (prozac).
00:32:00 – Štúdia ukazuje, že priame požitie glukorafanínu u myší je podobne účinné pri prevencii depresie zo sociálneho modelu stresu.
00:33:01 – Začiatok sekcie neurodegenerácie.
00:33:30 – Sulforafán a Alzheimerova choroba.
00:33:44 – Sulforaphane a Parkinsonova choroba.
00:33:51 - Sulforaphane a Hungtingtonova choroba.
00:34:13 – Sulforaphane zvyšuje proteíny tepelného šoku.
00:34:43 – Začiatok sekcie traumatického poranenia mozgu.
00:35:01 – Sulforaphane podaný ihneď po TBI zlepšuje pamäť (štúdia na myšiach).
00:35:55 – Sulforafán a plasticita neurónov.
00:36:32 – Sulforaphane zlepšuje učenie v modeli diabetu typu II u myší.
00:37:19 – Sulforaphane a Duchennova svalová dystrofia.
00:37:44 – Inhibícia myostatínu vo svalových satelitných bunkách (in vitro).
00:38:06 – Neskoré video rekapitulácia: úmrtnosť a rakovina, poškodenie DNA, oxidačný stres a zápal, vylučovanie benzénu, kardiovaskulárne ochorenia, diabetes typu II, účinky na mozog (depresia, autizmus, schizofrénia, neurodegenerácia), dráha NRF2.
00:40:27 – Myšlienky na to, ako zistiť dávku brokolicových klíčkov alebo sulforafanu.
00:41:01 – Anekdoty o klíčení doma.
00:43:14 – O teplotách varenia a aktivite sulforafanu.
00:43:45 – Premena sulforafanu z glukorafanínu črevnými baktériami.
00:44:24 – Doplnky fungujú lepšie v kombinácii s aktívnou myrozinázou zo zeleniny.
00:44:56 – Techniky varenia a krížová zelenina.
00:46:06 – Izotiokyanáty ako struma.
Závery
V súčasnosti sa veľa výskumov zameralo na úlohu signálnej dráhy Nrf2/Keap1/ARE pri zápale. Medzi enzýmami upregulovanými Nrf2 je HO-1 jedným z reprezentatívnych enzýmov stresovej reakcie. HO-1 má výrazné protizápalové a antioxidačné vlastnosti. Vo všeobecnosti signálna dráha Nrf2 negatívne reguluje aj cytokíny, faktory uvoľňujúce chemokíny, MMP a ďalšie zápalové mediátory produkciu COX-2 a iNOS, ktoré priamo alebo nepriamo ovplyvňujú príslušné dráhy NF-kB a MAPK a ďalšie siete kontrolujúce zápal. Predpokladá sa, že signálne dráhy Nrf2 a NF-kB interagujú, aby regulovali transkripciu alebo funkciu downstream cieľových proteínov. Potlačenie alebo inaktivácia transkripčnej aktivity sprostredkovanej NF-kB prostredníctvom Nrf2 sa s najväčšou pravdepodobnosťou vyskytuje v skorej fáze zápalu, pretože NF-kB reguluje de novo syntézu radu prozápalových mediátorov. Vo výskume však stále existujú určité obmedzenia, ako napríklad to, či existujú spojenia medzi Nrf2 a inými signálnymi dráhami, ako je JAK/STAT, význam súčasných aktivátorov Nrf2 odvodených z prírodných rastlinných zdrojov pri zápale a ako zlepšiť biologickú aktivitu a zlepšiť zacielenie týchto zlúčenín. Tie si vyžadujú ďalšie experimentálne overenie.
Okrem toho môže signálna dráha Nrf2 regulovať > 600 génov [163], z ktorých > 200 kóduje cytoprotektívne proteíny [164], ktoré sú tiež spojené so zápalom, rakovinou, neurodegeneratívnymi ochoreniami a inými závažnými ochoreniami [165]. Rastúce dôkazy naznačujú, že signálna dráha Nrf2 je pri mnohých rakovinách deregulovaná, čo vedie k aberantnej expresii génovej batérie závislej od Nrf2. Okrem toho zápal hrá hlavnú úlohu pri ochoreniach súvisiacich s oxidačným stresom, najmä pri rakovine. Aplikácia niekoľkých Nrf2 aktivátorov na potlačenie zápalu môže viesť k aberantnej expresii Nrf2 downstream génov, ktoré indukujú onkogenézu a rezistenciu na chemo a/alebo rádioterapiu. Preto môžu byť vyvinuté vysoko špecifické aktivátory Nrf2, aby sa minimalizovali jeho pleiotropné účinky. Niekoľko aktivátorov Nrf2 preukázalo významné zlepšenie protizápalových funkcií pri ochoreniach súvisiacich s oxidačným stresom. Najlepším príkladom aktivátora Nrf2 schváleného FDA a široko používaného na liečbu zápalových ochorení, ako je roztrúsená skleróza (MS), je dimetylfumarát. Tecfidera� (registrovaný názov dimetylfumarátu od spoločnosti Biogen) účinne používaná na liečbu recidivujúcich foriem roztrúsenej sklerózy u veľkého počtu pacientov (152). Účinnosť použitia aktivátorov Nrf2 na liečbu zápalových ochorení si však vyžaduje ďalšie overenie, aby sa predišlo škodlivým účinkom Nrf2. Preto by vývoj terapií na protizápalovú aktivitu sprostredkovanú Nrf2 mohol mať významný klinický dopad. Prebiehajúce štúdie signálnej dráhy Nrf2 na celom svete sa venujú vývoju vysoko cielených terapeutických činidiel na kontrolu symptómov zápalu a na prevenciu a liečbu rakoviny, ako aj neurodegeneratívnych a iných závažných ochorení.
Na záver, Nrf2 sníma úrovne oxidačného stresu v ľudskom tele a v konečnom dôsledku pomáha podporovať reguláciu antioxidačných a detoxikačných enzýmov a génov. Pretože chronický zápal spôsobený zvýšenou úrovňou oxidačného stresu je spojený s neurodegeneratívnymi ochoreniami, Nrf2 môže hrať zásadnú úlohu pri liečbe zdravotných problémov, ako je okrem iného Alzheimerova choroba. Rozsah našich informácií je obmedzený na chiropraktické a zdravotné problémy chrbtice. Ak chcete prediskutovať túto tému, neváhajte sa opýtať Dr. Jimeneza alebo nás kontaktujte na adrese�915-850-0900 .
Ďalšia téma diskusie: Zmiernenie bolesti kolena bez operácie
Bolesť kolena je dobre známym príznakom, ktorý sa môže vyskytnúť v dôsledku rôznych zranení a/alebo stavov kolena, vrátane�športové zranenia. Koleno je jedným z najzložitejších kĺbov v ľudskom tele, pretože je tvorené priesečníkom štyroch kostí, štyroch väzov, rôznych šliach, dvoch meniskov a chrupavky. Podľa Americkej akadémie rodinných lekárov medzi najčastejšie príčiny bolesti kolena patrí patelárna subluxácia, patelárna tendinitída alebo skokanovo koleno a Osgood-Schlatterova choroba. Hoci bolesť kolena sa s najväčšou pravdepodobnosťou vyskytuje u ľudí starších ako 60 rokov, bolesť kolena sa môže vyskytnúť aj u detí a dospievajúcich. Bolesť kolena sa môže liečiť doma podľa metód RICE, avšak vážne poranenia kolena môžu vyžadovať okamžitú lekársku pomoc vrátane chiropraktickej starostlivosti. �
Neurodegeneratívne ochorenia, ako je Alzheimerova choroba a Parkinsonova choroba, postihujú milióny jedincov na celom svete. Na liečbu symptómov niekoľkých neurodegeneratívnych ochorení sú dostupné rôzne možnosti liečby, hoci výsledky sú často obmedzené. Výskumné štúdie zistili, že oxidačný stres spôsobený vnútornými aj vonkajšími faktormi môže byť príčinou rozvoja neurodegeneratívnych ochorení. The transkripčný faktor, Nrf2, bolo zistené, že funguje ako hlavný obranný mechanizmus proti oxidačnému stresu. Účelom nižšie uvedeného článku je ukázať účinky Nrf2 o neurodegeneratívnych ochoreniach.
Modulácia proteostázy transkripčným faktorom NRF2
Neurodegeneratívne ochorenia sú spojené s akumuláciou špecifických proteínových agregátov, čo naznačuje úzke spojenie medzi poraneným mozgom a stratou proteostázy. Proteostáza označuje všetky procesy, ktorými bunky riadia množstvo a skladanie proteómu vďaka širokej sieti, ktorá integruje reguláciu signálnych dráh, génovú expresiu a systémy degradácie proteínov. Tento prehľad sa pokúša zhrnúť najdôležitejšie zistenia o transkripčnej modulácii proteostázy vyvolanej transkripčným faktorom NRF2 (jadrový faktor (odvodený z erytroidov 2) podobný 2). NRF2 sa klasicky považuje za hlavný regulátor antioxidačnej bunkovej odpovede, hoci sa v súčasnosti objavuje ako kľúčová zložka transdukčného mechanizmu na udržanie proteostázy. Ako budeme diskutovať, NRF2 by sa dal predstaviť ako centrum, ktoré zostavuje núdzové signály odvodené z akumulácie nesprávne zložených proteínov, aby sa vytvorila koordinovaná a trvalá transkripčná odpoveď. To sa dosahuje funkciami NRF2 súvisiacimi s kontrolou génov zapojených do udržiavania fyziológie endoplazmatického retikula, proteazómu a autofágie.
Nuclear Factor (erytroid-derived 2)-like 2 (NRF2) je základný leucínový zipsový proteín, ktorý sa v súčasnosti považuje za hlavný regulátor bunkovej homeostázy. Riadi bazálnu a stresom indukovateľnú expresiu viac ako 250 génov, ktoré majú spoločný cis-pôsobiaci zosilňovač nazývaný prvok antioxidačnej odozvy (ARE) [1], [2], [3], [4], [5]. Tieto gény sa podieľajú na detoxikačných reakciách fázy I, II a III, metabolizme glutatiónu a peroxiredoxínu/tioredoxínu, produkcii NADPH prostredníctvom pentózofosfátovej dráhy a jablčného enzýmu, oxidácii mastných kyselín, metabolizme železa a proteostáze [6]. Vzhľadom na tieto široké cytoprotektívne funkcie je možné, že jediný farmakologický zásah v NRF2 by mohol zmierniť účinok hlavných vinníkov chronických ochorení, vrátane oxidačného, zápalového a proteotoxického stresu. Úlohou NRF2 pri modulácii antioxidačnej obrany a riešení zápalu sa zaoberalo množstvo štúdií (prehľad v [7]). Tu sa zameriame na jeho úlohu v proteostáze, tj homeostatickej kontrole proteínovej syntézy, skladania, obchodovania a degradácie. Príklady budú poskytnuté v kontexte neurodegeneratívnych ochorení.
Strata proteostázy ovplyvňuje aktivitu NRF2 pri neurodegeneratívnych ochoreniach
Všeobecným znakom neurodegeneratívnych ochorení je výskyt aberantnej agregácie niektorých proteínov. Nesprávne poskladané proteínové agregáty a-synukleínu (a-SYN) sa teda nachádzajú v Parkinsonovej chorobe (PD), y-amyloidných (Ap) plakoch a hyperfosforylovaných TAU neurofibrilárnych spletiach pri Alzheimerovej chorobe (AD), huntingtín (Htt) v Huntingtonova choroba (HD), superoxiddismutáza 1 (SOD1) a TAR DNA viažuci proteín 43 (TDP-43) pri amyotrofickej laterálnej skleróze (ALS), priónový proteín (PrP) pri spongiformných encefalopatiách atď. Proteínové agregáty môžu mať vplyv na viaceré bunkových dráh, čo môže následne ovplyvniť hladiny a aktivitu NRF2.
Rôzne úrovne regulácie prísne kontrolujú aktivitu NRF2
Za fyziologických podmienok bunky vykazujú nízke hladiny proteínu NRF2 kvôli jeho rýchlemu obratu. V reakcii na rôzne stimuly sa hromadí proteín NRF2, vstupuje do jadra a zvyšuje transkripciu génov obsahujúcich ARE. Preto je riadenie hladín proteínu NRF2 kľúčovým bodom, ktorý by mal integrovať pozitívne a negatívne vstupné signály. Ako budeme ďalej diskutovať, NRF2 je aktivovaný rôznymi prekrývajúcimi sa mechanizmami na organizáciu rýchlej a efektívnej reakcie, ale na druhej strane by NRF2 mohol byť inhibovaný, pravdepodobne v druhej fáze, aby sa jeho reakcia vypla.
Z klasického hľadiska bola aktivácia NRF2 považovaná za dôsledok bunkovej odpovede na oxidačné alebo elektrofilné zlúčeniny. V tomto ohľade hrá kľúčovú úlohu adaptér ubikvitín E3 ligázy Kelch-like ECH-associated protein 1 (KEAP1). Molekulárnym detailom sa budeme ďalej venovať v časti 4.1. Stručne povedané, KEAP1 pôsobí ako redoxný senzor v dôsledku kritických cysteínových zvyškov, ktoré vedú k ubikvitinácii NRF2 a proteazomálnej degradácii. Okrem tejto klasickej modulácie je NRF2 hlboko regulovaný signalizáciou udalostí. V skutočnosti sa ukázalo, že rôzne kinázy fosforylujú a regulujú NRF2. Napríklad NRF2 môže byť fosforylovaný mitogénom aktivovanými proteínkinázami (MAPK), hoci jeho príspevok k aktivite NRF2 zostáva nejasný [8], [9], [10], [11]. PKA kináza, ako aj niektoré izoenzýmy PKC [12], CK2 [13] alebo Fyn [14] fosforylujú NRF2, čím modifikujú jeho stabilitu. Predchádzajúca práca našej skupiny uviedla, že glykogénsyntáza kinse-3? (GSK-3?) inhibuje NRF2 vylúčením jadra a proteazomálnou degradáciou [15], [25], [26], [27], [28], [29], [30]. Molekulárne detaily budú diskutované v časti 4.1. Okrem toho NRF2 podlieha iným typom regulácie. Napríklad acetylácia NRF2 pomocou CBP/p300 zvyšuje jeho aktivitu [17], zatiaľ čo je inhibovaná miR153, miR27a, miR142-5p a miR144 [16] alebo metyláciou ostrovčekov cytozín-guanín (CG) v promótore NRF2. [18].
Vplyv proteínových agregátov na regulačné mechanizmy NRF2
V tejto časti sa zameriame na to, ako by akumulácia nesprávne poskladaného proteínu mohla ovplyvniť aktivitu NRF2 a poskytnúť niektoré z vyššie uvedených dráh ako ilustratívne príklady. Po prvé, musíme vziať do úvahy, že akumulácia bielkovín bola úzko spojená s oxidačným poškodením. Akumulácia a agregácia nesprávne zložených proteínov skutočne vyvoláva abnormálnu produkciu reaktívnych foriem kyslíka (ROS) z mitochondrií a iných zdrojov [19]. Ako je uvedené vyššie, ROS bude modifikovať redox-senzitívne cysteíny KEAP1, čo vedie k uvoľneniu, stabilizácii a jadrovej lokalizácii NRF2.
Pokiaľ ide o proteinopatie, príkladom dysregulovaných signalizačných udalostí, ktoré môžu ovplyvniť NRF2, je hyperaktivácia GSK-3? v AD. GSK-3p, tiež známy ako TAU kináza, sa podieľa na fosforylácii tohto proteínu spojeného s mikrotubulami, čo vedie k jeho agregácii, tvorbe neurofibrilárnych spletí a prerušeniu axonálneho transportu (prehľad v [20]). Na druhej strane, GSK-3? dramaticky znižuje hladiny a aktivitu NRF2, ako je uvedené vyššie. Hoci to nie je široko akceptované, amyloidná kaskáda naznačuje, že toxický A? oligoméry zvyšujú GSK-3? aktivita spolu s hyperfosforyláciou TAU a smrťou neurónov [21], [22]. Existujú rôzne modely na vysvetlenie toho, ako A? uprednostňuje GSK3-? činnosť. Napríklad A? sa viaže na inzulínový receptor a inhibuje signálne dráhy PI3K a AKT, ktoré sú kľúčové pre udržanie GSK-3? inaktivovaný fosforyláciou na svojom N-terminálnom zvyšku Ser9 [23]. Na druhej strane, extracelulárny A? interaguje s Frizzled receptormi, blokuje WNT signalizáciu [24] a opäť vedie k uvoľneniu aktívneho GSK-3?. Stručne povedané, A? akumulácia vedie k abnormálnej hyperaktivácii GSK-3p, čím sa zhoršuje primeraná odpoveď NRF2.
Ako je uvedené v nasledujúcej časti, nesprávne poskladané proteíny vedú k aktivácii PERK a MAPK, ktoré následne upregulujú NRF2 [31], [8], [9], [10], [11]. Okrem toho bola pri niekoľkých proteinopatiách hlásená dysregulovaná aktivita CBP/p300 [32] a preukázal sa aj globálny pokles metylácie DNA v mozgoch AD [33], čo poskytuje dôvod na preskúmanie relevantnosti týchto zistení v regulácii NRF2.
My a iní sme pozorovali pri nekropsiách pacientov s PD a AD zvýšenie hladín proteínu NRF2 a niektorých jeho cieľov, ako je hemoxygenáza 1 (HMOX1), NADPH chinónoxidáza 1 (NQO1), p62 atď., a to imunoblotom a imunohistochémiou [34], [35], [36], [37], [38], [39]. Up-regulácia NRF2 pri týchto chorobách sa interpretuje ako neúspešný pokus chorého mozgu obnoviť homeostatické hodnoty. Iná štúdia však naznačila, že NRF2 je prevažne lokalizovaný v cytoplazme AD hipokampálnych neurónov, čo naznačuje zníženú transkripčnú aktivitu NRF2 v mozgu [40]. Je možné, že rozdiel v týchto pozorovaniach súvisí so zmenami faktorov, ktoré kontrolujú NRF2 v progresívnych štádiách neurodegenerácie.
K proteostáze prispievajú tri hlavné systémy, a to rozložená proteínová odpoveď (UPR), ubikvitínový proteazómový systém (UPS) a autofágia. Ďalej uvádzame dôkazy, že si NRF2 predstavujeme ako rozbočovač spájajúci núdzové signály aktivované proteínovými agregátmi s mechanizmom proteínových derivátov.
NRF2 sa zúčastňuje na roztvorenej proteínovej odpovedi (UPR)
Aktivácia NRF2 v reakcii na UPR
Oxidačné skladanie proteínov v ER je poháňané množstvom odlišných dráh, z ktorých najkonzervovanejšia zahŕňa proteín disulfid-izomerázu (PDI) a sulfhydryloxidázový endoplazmatický oxidoreduktín 1 (ERO1p a ERO1p u cicavcov) ako donor disulfidu. Stručne povedané, PDI katalyzuje tvorbu a štiepenie disulfidových väzieb medzi cysteínovými zvyškami v proteínoch, keď sa skladajú, v dôsledku redukcie a oxidácie vlastných cysteínových aminokyselín. PDI sa recykluje pôsobením upratovacieho enzýmu ERO1, ktorý znovu zavádza disulfidové väzby do PDI [41]. Molekulárny kyslík je terminálnym akceptorom elektrónov ERO1, ktorý generuje stechiometrické množstvá peroxidu vodíka na každú vytvorenú disulfidovú väzbu [42]. Peroxidázy (PRX4) a glutatiónperoxidázy (GPX7 a GPX8) sú kľúčové enzýmy na zníženie peroxidu vodíka v ER. Keď tento oxidoredukčný systém nefunguje správne, dochádza v ER k abnormálnej akumulácii nesprávne poskladaných proteínov a do cytoplazmy a jadra sa prenesie súbor signálov nazývaných odpoveď na rozbalený proteín (UPR), aby sa obnovila homeostáza ER [43]. Na snímanie stresu ER u eukaryotov boli identifikované tri proteíny spojené s membránou: aktivačný transkripčný faktor 6 (ATF6), pankreatický ER eIF2? kináza (PERK, tiež dvojvláknová RNA-aktivovaná proteínkináza podobná ER kináze) a kináza 1 vyžadujúca inozitol (IRE1). Luminálna doména každého senzora je naviazaná na 78 kDa chaperón nazývaný glukózou regulovaný proteín (GRP78/BIP). BIP disociuje po strese ER, aby naviazal nezbalené proteíny, čo vedie k aktivácii troch senzorov [44].
NRF2 a jeho homológ NRF1, tiež súvisiaci s antioxidačnou odpoveďou, sa podieľajú na transdukcii UPR do jadra. V prípade NRF1 sa tento proteín nachádza na membráne ER a po deglykozylácii alebo štiepení podlieha jadrovej translokácii. Potom aktivácia UPR vedie k spracovaniu NRF1 a jadrovej akumulácii výsledného fragmentu v jadrovom kompartmente. Schopnosť transaktivovať gény tohto fragmentu NRF1 obsahujúce ARE je však stále predmetom diskusie [45].
Glover-Cutter a spolupracovníci preukázali aktiváciu ortológu NRF2 C. elegans, SKN-1, s rôznymi ER stresormi. Zvýšená expresia SKN-1 bola závislá od rôznych mediátorov UPR, vrátane ortológov červov IRE1 alebo PERK [46]. V bunkách s deficitom PERK vedie narušená syntéza proteínov k akumulácii endogénnych peroxidov a následnej apoptóze [47]. Efektor používaný PERK na ochranu ER pred týmito peroxidmi môže byť NRF2, pretože sa uvádza, že PERK fosforyluje NRF2 na Ser40, čím bráni jeho degradácii KEAP1 [31]. Indukcia ASK1 pravdepodobne zohráva úlohu aj v tejto ceste prostredníctvom kinázového účinku IRE2 sprostredkovaného TRAF1 [48]. Hoci úloha MAPK pri regulácii NRF2 je stále kontroverzná, nedávno sa navrhlo, že dráha IRE1-TRAF2-ASK1-JNK môže aktivovať NRF2 [49] (obr. 1). Je zaujímavé, že v C. elegans a ľudských bunkách nové dôkazy naznačujú, že cysteínová sulfenylácia kinázy IRE1 v jej aktivačnej slučke inhibuje UPR sprostredkovanú IRE1 a iniciuje antioxidačnú odpoveď p38 riadenú NRF2. Údaje naznačujú, že IRE1 má starodávnu funkciu ako cytoplazmatický sentinel, ktorý aktivuje p38 a NRF2 [50].
Obrázok 1 Regulácia NRF2 UPR. Akumulácia neposkladaných alebo nesprávne poskladaných proteínov vo vnútri endoplazmatického retikula môže iniciovať odpoveď na rozbalený proteín (UPR). Najprv sa chaperón BIP uvoľní z intraluminálnej domény ER senzorov IRE1 a PERK, aby sa naviazali nerozbalené/chybne poskladané proteíny. To umožňuje dimerizáciu a trans-autofosforyláciu ich cytosolických domén. Aktivácia PERK vedie k priamej fosforylácii NRF2 na Ser40, čo vedie k translokácii NRF2 do jadra a aktivácii cieľových génov. Aktivácia IRE1 indukuje nábor TRAF2, po ktorom nasleduje fosforylácia a aktivácia ASK1 a JNK. Keďže sa uvádza, že JNK fosforyluje a aktivuje NRF2, je rozumné si myslieť, že aktivácia IRE1 by viedla k zvýšenej aktivite NRF2.
Mnoho štúdií o indukcii UPR sa uskutočnilo s inhibítorom glykozylácie proteínov tunicamycínom. Zdá sa, že NRF2 je nevyhnutný na prevenciu apoptotickej bunkovej smrti vyvolanej tunicamycínom [31] a jeho aktivácia za týchto podmienok je riadená autofagickou degradáciou KEAP1 [51]. V súlade s tým shRNA-sprostredkované umlčanie expresie NRF2 v bunkách aTC-6, línii myších buniek inzulinómu, významne zvýšilo cytotoxicitu indukovanú tunicamycínom a viedlo k zvýšeniu expresie proapoptotického stresového markera ER CHOP10. Na druhej strane aktivácia NRF2 pomocou 1,2-ditiol-3-tiónu (D3T) znížila cytotoxicitu tunikamycínu a zoslabila expresiu CHOP10 a PERK [52]. Je zaujímavé, že čuchové neuróny podrobené systémovej aplikácii tunicamycínu zvýšili NRF2 paralelne s ostatnými členmi UPR, ako sú CHOP, BIP, XBP1 [53]. Tieto výsledky boli rozšírené na štúdie in vivo, pretože laterálna ventrikulárna infúzia tunicamycínu u potkanov vyvolala expresiu PERK a NRF2 v hipokampe sprevádzanú významnými kognitívnymi deficitmi, zvýšenou fosforyláciou TAU a depozitmi Ap42 [54].
NRF2 Up-reguluje kľúčové gény pre udržanie fyziológie ER
Lumen ER potrebuje dostatok GSH z cytosolu, aby sa zachovala disulfidová chémia. NRF2 moduluje kľúčové enzýmy metabolizmu GSH v mozgu, ako je transport cystínu/glutamátu, β-glutamátcysteínsyntetáza (a-GS), katalytické a modulátorové podjednotky glutamát-cysteínligázy (GCLC a GCLM), glutatiónreduktáza (GR) a glutatiónperoxidáza (GPX) (prehľad v [55]). Význam NRF2 pri udržiavaní GSH v ER je podporený zistením, že farmakologická alebo genetická aktivácia NRF2 vedie k zvýšenej syntéze GSH prostredníctvom GCLC/GCLM, zatiaľ čo inhibícia expresie týchto enzýmov pomocou NRF2-knockdown spôsobila akumuláciu poškodených buniek. proteínov v ER, čo vedie k aktivácii UPR [56].
V C. elegans niekoľko komponentov cieľových génov UPR regulovaných SKN-1, vrátane Ire1, Xbp1 a Atf6. Hoci NRF2 up-reguluje expresiu niekoľkých génov pre peroxidázu (PRX) a glutatiónperoxidázu (GPX) u cicavcov (prehľad v [57]), iba GPX8 je bona fide ER-lokalizovaný enzým, ktorý prechováva vyhľadávací signál KDEL [58]. Strata GPX8 spôsobuje aktiváciu UPR, únik peroxidu vodíka odvodeného od ERO1a do cytosolu a bunkovú smrť. Peroxid vodíka odvodený od ERO1? aktivita nemôže difundovať z ER do cytosolu v dôsledku spoločného pôsobenia GPX8 a PRX4 [59]. V tomto ohľade analýza expresného poľa génovej dráhy antioxidačnej obrany s použitím RNA z tkaniva myší divokého typu a tkaniva s nulovou hodnotou NRF2 odhalila, že expresia GPX8 bola znížená v neprítomnosti NRF2 [60]. V súlade s tým, transkriptómová analýza zo vzoriek pacientov trpiacich myeloproliferatívnymi novotvarmi, polycytémiou alebo myelofibrózou, ochoreniami tiež spojenými s oxidačným stresom a chronickým zápalom nízkeho stupňa, ukazuje nižšie hladiny expresie NRF2 aj GPX8 v porovnaní s kontrolnými subjektmi [61]. Zatiaľ neexistujú štúdie, ktoré by špecificky zapájali GPX8 do ochrany ľudského mozgu, ale analýza transkriptómu u myší naznačuje kompenzačné zvýšenie GPX8 v reakcii na parkinsonský toxín MPTP [62].
Vplyv NRF2 na dysreguláciu UPR pri neurodegeneratívnych ochoreniach
Nesprávna funkcia enzýmov PDI a chronická aktivácia UPR môže následne iniciovať alebo urýchliť neurodegeneráciu. Neuróny postihnuté chorobou, zvieracie modely neurodegeneratívneho ochorenia, ako aj ľudské tkanivá po smrti, preukázali up-reguláciu niekoľkých UPR-markerov vo väčšine týchto porúch. Zmena dráhy PDI/UPR pri neurodegeneratívnych ochoreniach bola pekne zhrnutá v [63], ale mali by sa vziať do úvahy nasledujúce zvýraznenia z posmrtných vzoriek mozgu. Hladiny PDI sú zvýšené v neurónoch nesúcich spleť a v Lewyho telieskach u pacientov s AD a PD [64], [65]. PDI a ERP57 sú up-regulované v CSF od pacientov s ALS a v mozgoch pacientov s CJD [66], [67], [68]. BIP, PERK, IRE1 a ATF6 sú zvýšené vo vzorkách od pacientov s AD, PD alebo ALS [69], [70], [71], [67]. BIP, CHOP a XBP1 sú zvýšené v postmortálnych vzorkách mozgu z HD [72], [73]. Okrem toho sa v tkanivách mozgovej kôry u pacientov s CJD zistila up-regulácia ERP57, GRP94 a BIP [74]. Celkovo tieto dôkazy odhaľujú, že akumulácia nesprávne poskladaných proteínov v mozgovom parenchýme vedie k škodlivej a chronickej aktivácii UPR. Je zaujímavé, že existuje nedávna štúdia spájajúca aktiváciu NRF2 pomocou PERK na začiatku AD. V tejto štúdii autori analyzovali, či zmeny sprostredkované oxidačným stresom v NRF2 a UPR môžu predstavovať skoré udalosti v patogenéze AD pomocou ľudských buniek periférnej krvi a modelu transgénnej myši AD v rôznych štádiách ochorenia. Zvýšený oxidačný stres a zvýšený pSer40-NRF2 boli pozorované v ľudských mononukleárnych bunkách periférnej krvi izolovaných od jedincov s miernou kognitívnou poruchou. Okrem toho hlásili zhoršenú ER kalciovú homeostázu a up-regulované ER-stresové markery v týchto bunkách od jedincov s miernou kognitívnou poruchou a miernou AD [75].
Vzájomná regulácia NRF2 a ubikvitínového proteazómového systému (UPS)
UPS moduluje hladiny proteínu NRF2
UPS sa podieľa na degradácii poškodených alebo nesprávne poskladaných proteínov a riadi hladiny kľúčových regulačných molekúl v cytosóle a jadre. Centrálnym jadrom tohto systému je veľký multipodjednotkový enzým, ktorý obsahuje proteolyticky aktívny komplex s názvom 20S. Jadrový proteazóm 20S degraduje nezbalené proteíny, ale väzba na rôzne komplexy regulačných proteínov mení jeho substrátovú špecifickosť a aktivitu. Napríklad pridanie jednej alebo dvoch regulačných podjednotiek 19S k jadru 20S tvorí proteazóm 26S a mení jeho špecifickosť voči natívnym zloženým proteínom [76], [77]. Proteazomálna degradácia vyžaduje kovalentnú väzbu ubikvitínu. Konjugácia ubikvitínu prebieha prostredníctvom trojstupňového kaskádového mechanizmu. Po prvé, enzým aktivujúci ubikvitín E1 aktivuje ubikvitín v reakcii vyžadujúcej ATP. Potom jeden enzým E2 (proteín nosiča ubikvitínu alebo enzým konjugujúci ubikvitín) prenesie aktivovaný ubikvitín z E1 do substrátu, ktorý je špecificky viazaný na člena rodiny ubikvitín-proteín ligázy s názvom E3. Hoci presný osud ubikvitinovaného proteínu bude závisieť od povahy ubikvitínového reťazca, tento proces vo všeobecnosti vedie k degradácii proteazómom 26S [78].
E3-ligáza KEAP1 je najznámejším inhibítorom NRF2. Mechanizmus regulácie KEAP1 elegantne vysvetľuje, ako sa hladiny NRF2 prispôsobujú výkyvom oxidantov. Za základných podmienok je novo syntetizovaný NRF2 zachytený homodimérom KEAP1, ktorý viaže jednu molekulu NRF2 na dve aminokyselinové sekvencie s nízkou (aspartát, leucín, glycín; DLG) a vysokou (glutamát, treonín, glycín, glutamát; ETGE) afinitou. Interakcia s KEAP1 pomáha prezentovať NRF2 proteínovému komplexu CULLIN3/RBX1, čo vedie k jeho ubikvitinácii a následnej proteazomálnej degradácii. Redoxná modifikácia KEAP1 však bráni prezentácii NRF2 do UPS reprezentovaného CULLIN3/RBX1. Výsledkom je, že novosyntetizovaný NRF2 uniká degradácii závislej od KEAP1, hromadí sa v jadre a aktivuje gény obsahujúce ARE [79], [80], [81], [82].
E3-ligázový adaptér a-TrCP je tiež homodimér, ktorý sa zúčastňuje na signalizačných udalostiach súvisiacich s fosforyláciou NRF2 pomocou GSK-3p. Táto kináza fosforyluje špecifické serínové zvyšky NRF2 (aspartát, serín, glycín, izoleucín serín; DSGIS), aby sa vytvorila degradačná doména, ktorú potom rozpozná a-TrCP a označí ju na degradáciu proteazómu komplexom CULLIN1/RBX1. Identifikácia špecifických aminokyselín, ktoré sú fosforylované GSK-3? v tomto degrone sa uskutočnila kombinácia miestne cielenej mutagenézy Neh6 domény, 2D-gélovej elektroforézy [15], [26] a hmotnostnej spektroskopie [83]. V dôsledku toho inhibícia GSK-3? vysoko selektívnymi liekmi alebo siRNA proti izoformám GSK-3 viedlo k zvýšeniu hladín proteínu NRF2. Podobné výsledky sa našli so siRNA proti izoformám a-TrCP 1 a 2. Stabilizácia NRF2 po GSK-3? k inhibícii došlo v KEAP1-deficientných myších embryonálnych fibroblastoch a v ektopicky exprimovanom mutante s deléciou NRF2, ktorému chýbajú kritické zvyšky ETGE pre vysokoafinitnú väzbu na KEAP1, čo ďalej demonštruje reguláciu nezávislú od KEAP1.
V kontexte neurodegeneratívnych ochorení si môžeme predstaviť moduláciu NRF2 pomocou UPS dvoma rôznymi spôsobmi. Na jednej strane by systém KEAP1 vnímal redoxnú nerovnováhu odvodenú od nesprávne poskladanej akumulácie proteínov, zatiaľ čo os GSK-3/a-TrCP by pôsobila ako aktívny účastník signalizačnej transdukcie zmenenej stratou proteostázy (obr. 2).
Obrázok 2 UPS prísne kontroluje hladiny NRF2. V homeostatických podmienkach sú nízke hladiny NRF2 udržiavané pôsobením E3 ligázových adaptérov KEAP1 a a-TrCP. Vľavo sa NRF2 viaže na Kelch domény homodiméru KEAP1 prostredníctvom motívov s nízkou (DLG) a vysokou (ETGE) afinitou. Prostredníctvom svojej domény BTB sa KEAP1 súčasne viaže na komplex CULLIN3 / RBX1, čo umožňuje ubikvitináciu a degradáciu NRF2 proteazómom 26 S. Navyše GSK-3? fosforyluje Ser335 a Ser338 zvyšky NRF2, aby vytvorila degradačnú doménu (DpSGIpSL), ktorá je potom rozpoznávaná adaptérom ubikvitín ligázy a-TrCP a označená na degradáciu proteazómu komplexom CULLIN3/RBX1. Správne, po vystavení reaktívnym druhom kyslíka alebo elektrofilom sa kritické zvyšky Cys v KEAP1 upravia, čím sa KEAP1 stane neschopným účinne interagovať s NRF2 alebo CULLIN3/RBX1 a potom tento transkripčný faktor zvyšuje svoj polčas a transkripčnú aktivitu voči génom ARE. Signálne dráhy, ktoré vedú k inhibícii GSK-3a, ako je fosforylácia AKT na Ser9, majú za následok zhoršenú degradáciu NRF2 proteazómom, akumuláciu a indukciu cieľových génov.
NRF2 zvyšuje aktivitu UPS prostredníctvom kontroly transkripcie proteazómových podjednotiek
NRF2 up-reguluje expresiu niekoľkých proteazómových podjednotiek, čím chráni bunku pred akumuláciou toxických proteínov. Zdá sa, že dvadsať génov súvisiacich s proteazómom a ubikvitináciou je regulovaných NRF2, podľa širokej mikročipovej analýzy z pečeňovej RNA, ktorá bola nastavená pomocou NRF2 induktora D3T [84]. V ďalšej štúdii tí istí autori dokázali, že expresia väčšiny podjednotiek proteazómu 26S bola zvýšená až trojnásobne v pečeni myší liečených D3T. Hladiny podjednotkového proteínu a aktivita proteazómu boli koordinovane zvýšené. U myší, kde bol transkripčný faktor NRF2 narušený, však nebola pozorovaná žiadna indukcia. Aktivita promótora proteazómovej podjednotky PSMB5 (20S) sa zvýšila buď nadmernou expresiou NRF2, alebo liečbou aktivátormi v myších embryonálnych fibroblastoch a ARE boli identifikované v proximálnom promótore PSMB5 [85]. Farmakologická aktivácia NRF2 viedla k zvýšeným hladinám expresie reprezentatívnych proteazómových podjednotiek (PSMA3, PSMA6, PSMB1 a PSMB5) len v nesenescentných ľudských fibroblastoch obsahujúcich funkčný NRF2 [86]. Aktivácia NRF2 počas adaptácie na oxidačný stres vedie k vysokej expresii PSMB1 (20S) a PA28? podjednotky (alebo S11, regulátor proteazómu) [87]. Okrem toho výsledky z ľudských embryonálnych kmeňových buniek odhalili, že NRF2 riadi expresiu proteazómového maturačného proteínu (POMP), proteazómového chaperónu, ktorý zasa moduluje proliferáciu samoobnovujúcich sa ľudských embryonálnych kmeňových buniek, diferenciáciu troch zárodočných vrstiev a bunkové preprogramovanie [ 88]. Všetky tieto štúdie naznačujú, že NRF2 up-reguluje expresiu kľúčových komponentov UPS, a preto aktívne prispieva k odstraňovaniu proteínov, ktoré by inak boli toxické.
Os NRF2-UPS pri neurodegeneratívnych ochoreniach
Úloha UPS pri neurodegeneratívnych ochoreniach je oblasťou intenzívnych diskusií. Počiatočné štúdie uvádzali zníženú aktivitu proteazómu pri ľudských nekropsiách pacientov postihnutých niekoľkými neurodegeneratívnymi ochoreniami. Iné štúdie využívajúce prístupy in vitro a in vivo však zistili nezmenenú alebo dokonca zvýšenú aktivitu proteazómu (prehľad v [89]). Jedným z možných vysvetlení tejto nezrovnalosti je, že hladiny komponentov UPS sa môžu meniť počas progresie ochorenia a v rôznych oblastiach mozgu, ako to bolo navrhnuté pre ciele NRF2.
Napriek tejto kontroverzii je potrebné poznamenať, že up-regulácia proteazómových génov obsahujúcich ARE posilní UPS zvýšením klírensu toxických proteínov v mozgu. V skutočnosti ablácia NRF1, tiež modulátora antioxidačnej odpovede, v neurónových bunkách vedie k zhoršenej proteazómovej aktivite a neurodegenerácii. Experimenty s imunoprecipitáciou chromatínu a transkripčná analýza ukázali, že PSMB6 je regulovaný NRF1. Okrem toho profilovanie génovej expresie viedlo k identifikácii NRF1 ako kľúčového transkripčného regulátora proteazómových génov v neurónoch, čo naznačuje, že poruchy v NRF1 môžu prispievať k patogenéze neurodegeneratívnych ochorení [90]. Je zaujímavé, že sa ukázalo, že NRF1 a jeho dlhá izoforma nazývaná TCF11 up-regulujú proteazómové gény obsahujúce ARE po inhibícii proteazómu v spätnoväzbovej slučke, aby kompenzovali zníženú proteolytickú aktivitu [91], [92].
Čo sa týka NRF2, existuje korelácia medzi znížením hladín NRF2, RPT6 (19 S) a PSMB5 (20 S) v strednom mozgu myší s deficitom DJ-1 liečených neurotoxínom paraquatom [93]. Okrem toho prirodzene sa vyskytujúca zlúčenina sulforafan (SFN) poskytuje robustnejší obraz NRF2 ako kľúčového modulátora UPS. In vitro experimenty s myšacími neuroblastómovými bunkami Neuro2A preukázali zvýšenú expresiu katalytických podjednotiek proteazómu, ako aj jeho peptidázové aktivity v reakcii na SFN. Tento liek chránil bunky pred cytotoxicitou sprostredkovanou peroxidom vodíka a oxidáciou proteínov spôsobom závislým od funkcie proteazómu [94]. Okrem toho Liu a spolupracovníci použili reportérsku myš na monitorovanie aktivity UPS v reakcii na SFN v mozgu. Tieto myši všadeprítomne exprimujú proteín zelenej fluorescencie (GFP) fúzovaný s konštitutívnym degradačným signálom, ktorý podporuje jeho rýchlu degradáciu pomocou UPS (GFPu). V mozgovej kôre SFN znížila hladinu GFPu s paralelným zvýšením aktivít podobných chymotrypsínu (PSMB5), kaspáze (PSMB2) a trypsínu (PSMB1) proteazómu 20S. Okrem toho ošetrenie buniek odvodených od Huntingtona pomocou SFN odhalilo, že aktivácia NRF2 zvýšila degradáciu mHtt a znížila cytotoxicitu mHtt [95]. Hlavným mechanizmom účinku SFN je indukcia NRF2 [96]. Špecifický príspevok NRF2 by sa mal riešiť využitím systémov NRF2-null v ďalších štúdiách.
Funkčné spojenie medzi NRF2 a makroautofágiou
Hladiny proteínu NRF2 sú modulované adaptérom Protein P62
Autofágia sa týka degradácie cytosolických zložiek vo vnútri lyzozómov. Tento proces sa používa na odstraňovanie dlhovekých a nesprávne poskladaných proteínov, ako aj poškodených organel. Priama súvislosť medzi NRF2 a autofágiou bola prvýkrát pozorovaná v súvislosti s adaptorovým proteínom p62, tiež nazývaným SQSTM1 [97], [98], [99], [100], [101]. Tento proteín premiestňuje ubikvitinované proteíny do proteazomálnych a lyzozomálnych degradačných strojov a sekvestruje poškodené proteíny do agregátov pred ich degradáciou. P62 predstavuje doménu spojenú s ubikvitínom (UBA) na väzbu na ubikvitinované proteíny a oblasť interagujúcu s LC3 (LIR) na integráciu s autofagozomálnou membránou prostredníctvom autofagického receptora LC3.
Hoci p62-sprostredkovaná indukcia NRF2 a jeho cieľových génov bola prvýkrát opísaná v roku 2007 [102], molekulárny mechanizmus nebol úplne pochopený, kým sa neobjavila jeho interakcia s KEAP1 [103], [98], [99], [100 ], [101]. Komatsu a spolupracovníci identifikovali oblasť interakcie KEAP1 (KIR) v p62, ktorá viazala KEAP1 v rovnakom základnom povrchovom vrecku ako NRF2 a s väzbovou afinitou podobnou motívu ETGE v NRF2, čo naznačuje konkurenciu medzi p62 a NRF2. Ukázalo sa, že fosforylácia Ser351 v KIR motíve v p62 (349-DPSTGE-354) zvyšuje jeho afinitu pre KEAP1, súťaží s väzbou NRF2 a umožňuje jeho akumuláciu a transkripčnú aktiváciu jeho cieľových génov [98], [99]. V skutočnosti nadmerná expresia p62 viedla k zníženej ubikvitinácii NRF2 a následnej stabilizácii, ako aj k indukcii jeho cieľových génov [104]. Predpokladá sa, že niektoré kinázy sa podieľajú na fosforylácii p62. Cicavčí cieľ rapamycínového komplexu 1 (mTORC1) môže byť zahrnutý, pretože liečba inhibítorom mTOR rapamycínom potlačila fosforyláciu p62 a down-reguláciu KEAP1 po liečbe arzenitom. Nedávno sa ukázalo, že kináza 1 aktivovaná TGF-a (TAK1) môže tiež fosforylovať p62, čím sa zvyšuje degradácia KEAP1 a up-regulácia NRF2. Autori tejto štúdie naznačujú, že je to spôsob, ako regulovať bunkovú redoxtázu v podmienkach ustáleného stavu, pretože nedostatok TAK1 zvyšuje reguláciu ROS v neprítomnosti akéhokoľvek exogénneho oxidantu v rôznych tkanivách myší paralelne so znížením hladín proteínu NRF2 [105 ].
Konštrukt p62 bez domény UBA bol stále schopný viazať KEAP1, čo znamená, že interakcia nezávisela od ubikvitinovaného KEAP1 [101]. Homológ p62 v Drosophila melanogaster, pomenovaný Ref (2), však neobsahuje motív KIR a priamo neinteraguje s DmKEAP1, hoci sa môže viazať na ubikvitinovaný DmKEAP1 prostredníctvom domény UBA. Okrem toho môže DmKEAP1 priamo interagovať s Atg8 (homológ s cicavčím LC3). Nedostatok KEAP1 vedie k indukcii Atg8 a autofágie v závislosti od ortológu CncC NRF2 a nezávisle od TFEB / MITF [106]. Zdá sa však, že vzťah medzi NRF2 a autofágiou je zachovaný, čo zdôrazňuje jeho funkčný význam.
Indukcia NRF2 pomocou p62 je výsledkom súťaže o väzbu KEAP1 a degradácie KEAP1 v lyzozóme. Umlčanie p62 pomocou siRNA zdvojnásobilo polčas KEAP1 paralelne so znížením NRF2 a jeho cieľových génov [101]. V súlade s tým, ablácia expresie p62 preukázala zvýšené hladiny KEAP1 v porovnaní s myšami divokého typu. Veľmi dôležité je, že prírastok hladín KEAP1 nebol ovplyvnený inhibítormi proteazómu, ale bol znížený pri autofágii vyvolávajúcej hladovanie [107]. V skutočnosti je KEAP1 prítomný v cicavčích bunkách v autofagických vezikulách zdobených p62 a LC3 [99], [100], [103]. Všetky tieto údaje naznačujú, že KEAP1 je substrátom makroautofágneho aparátu, ale tento problém by sa mal analyzovať podrobnejšie z dôvodu existencie niektorých kontroverzných výsledkov. Hladiny proteínu KEAP1 boli zvýšené u myší s nulou Atg7, kľúčového efektora makroautofágie [107], ale farmakologická inhibícia makroautofágie torínom1, E64/pepstatínom alebo bafilomycínom nedokázala akumulovať KEAP1 [107], [100]. Celkovo tieto výsledky naznačujú, že zvýšené hladiny p62 sekvestrujú KEAP1 do autofagických vakuol a pravdepodobne to vedie k autofagickej degradácii KEAP1 umožňujúcej aktiváciu NRF2 (obr. 3). Dve rôzne štúdie uvádzajú, že v tomto kontexte hrajú dôležitú úlohu reduktázy kyseliny sulfínovej SESTRINS. SESTRIN 2 interaguje s p62, KEAP1 a RBX1 a uľahčuje p62-dependentnú degradáciu KEAP1 a NRF2 aktiváciu cieľových génov [108]. Ďalšia štúdia ukázala, že SESTRIN 2 interagoval s ULK1 a p62, čím podporoval fosforyláciu p62 na Ser403, čo uľahčilo degradáciu nákladných proteínov vrátane KEAP1 [109].
Obrázok 3 Hladiny NRF2 sú regulované adaptorovým proteínom p62. Fosforylácia Ser 351 v KIR motíve p62 (349-DPSTGE-354) mTORC1, TAK1 alebo inými kinázami vedie k zvýšenej afinite k väzbe na KEAP1 v dôsledku podobnosti s motívom ETGE v NRF2. V dôsledku toho fosforylovaný p62 vytesňuje NRF2 a viaže KEAP1. Motív LIR v p62 umožňuje interakciu s LC3 v autofagozomálnej membráne, takže komplex p62-KEAP1 je nakoniec degradovaný v lyzozóme. V dôsledku toho je NRF2 schopný akumulovať sa, premiestňovať sa do jadra a zvyšovať transkripciu génov obsahujúcich ARE, vrátane p62. Tento regulačný mechanizmus poskytuje trvalú odpoveď NRF2, pretože KEAP1 musí byť novo syntetizovaný, aby sa inhibovala aktivita NRF2.
Modulácia makroautofágových génov pomocou NRF2
NRF2 reguluje expresiu relevantných génov pre makroautofágiu, ako aj pre UPR a UPS. Prvý dôkaz pochádza zo štúdií, v ktorých sa ukázalo, že expresia p62 je indukovaná po expozícii elektrofilom, ROS a oxidu dusnatému [110], [111], [112]. Mechanizmus indukcie bol opísaný o niekoľko rokov neskôr so zistením, že p62 obsahuje vo svojom génovom promótore funkčný ARE [99]. V nedávnej štúdii sa po bioinformatickej analýze a testoch ChIP našlo a overilo niekoľko ďalších funkčných ARE. Okrem toho myšie embryonálne fibroblasty a kortikálne neuróny z myší s knockoutom Nrf2 vykazovali zníženú expresiu p62, ktorú bolo možné zachrániť lentivírusom exprimujúcim NRF2. Podobne nedostatok NRF2 znížil hladiny p62 v poranených neurónoch z myšacieho hipokampu [36]. Preto sa navrhlo, že aktivácia NRF2 zvyšuje hladiny p62, čo vedie k degradácii KEAP1 a uprednostňuje ďalšiu stabilizáciu NRF2 v slučke pozitívnej spätnej väzby. Tento nekanonický mechanizmus indukcie NRF2 vyžaduje zmeny v génovej expresii a môže byť relevantnou odpoveďou na predĺžený bunkový stres.
Ukázalo sa, že proteín rozpoznávajúci náklad NDP52 je transkripčne regulovaný pomocou NRF2. NDP52 funguje podobným spôsobom ako p62, rozpoznáva ubikvitinované proteíny a interaguje s LC3 prostredníctvom domény LIR, takže náklad je degradovaný v lyzozómoch. V sekvencii DNA promótora Ndp52 sa našlo päť predpokladaných ARE. Tri z nich boli identifikované pomocou rôznych mutantných konštruktov a testov ChIP ako nevyhnutné pre transkripciu Ndp2 sprostredkovanú NRF52 [113]. Je potrebné poznamenať, že hladiny mRNA Ndp52 boli znížené v hipokampe myší s knockoutom Nrf2. Jedna z týchto sekvencií bola tiež validovaná v nezávislej štúdii ako NRF2-regulovaný ARE [36].
Úloha NRF2 pri modulácii autofágie však nie je obmedzená na indukciu týchto dvoch proteínov na rozpoznávanie nákladu. S cieľom získať hlbší prehľad o úlohe NRF2 pri modulácii ďalších génov súvisiacich s autofágiou naša skupina skrínovala databázu imunoprecipitácie chromatínu ENCODE na dva proteíny, MAFK a BACH1, ktoré viažu ARE regulované NRF2. Pomocou skriptu vygenerovaného z konsenzuálnej sekvencie ARE JASPAR sme identifikovali niekoľko domnelých ARE v mnohých autofágnych génoch. Dvanásť z týchto sekvencií bolo validovaných ako ARE regulované NRF2 v deviatich autofágových génoch, ktorých expresia bola znížená vo fibroblastoch myších embryí myší s knockoutom Nrf2, ale mohla byť obnovená lentivírusom exprimujúcim NRF2. Naša štúdia preukázala, že NRF2 aktivuje expresiu niektorých génov zapojených do rôznych krokov autofagického procesu, vrátane iniciácie autofágie (ULK1), rozpoznávania nákladu (p62 a NDP52), tvorby autofagozómov (ATG4D, ATG7 a GABARAPL1), predlžovania (ATG2B a ATG5 ) a autolyzozómový klírens (ATG4D). V dôsledku toho bol tok autofágy v reakcii na peroxid vodíka narušený, keď chýbal NRF2 [36].
Význam expresie makroautofágových génov sprostredkovaných NRF2 pri neurodegeneratívnych poruchách
Ukázalo sa, že defektná autofágia hrá dôležitú úlohu pri niekoľkých neurodegeneratívnych ochoreniach [114] a ablácia autofágie vedie k neurodegenerácii u myší [115], [116]. Myši s knockoutom Atg7 odhalili, že nedostatok autofágie vedie k akumulácii p62 v ubikvitín-pozitívnych inklúznych telieskach. KEAP1 bol sekvestrovaný v týchto inklúznych telieskach, čo viedlo k stabilizácii NRF2 a indukcii cieľových génov [103]. Dôležité je, že nadmerná akumulácia p62 spolu s ubikvitinovanými proteínmi bola identifikovaná pri neurodegeneratívnych ochoreniach, vrátane AD, PD a ALS [117]. V skutočnosti neuróny exprimujúce vysoké hladiny APP alebo TAU pacientov s AD tiež exprimovali p62 a jadrový NRF2, čo naznačuje ich pokus o degradáciu intraneuronálnych agregátov prostredníctvom autofágie [36].
Nedostatok NRF2 zhoršuje agregáciu proteínov v kontexte AD. V skutočnosti sa u myší s knockoutom Nrf2 nachádzajú zvýšené hladiny fosforylovaných a v sarkozyle nerozpustných TAU, hoci v porovnaní s pozadím divokého typu nebolo možné zistiť žiadny rozdiel v aktivitách kinázy alebo fosfatázy [113]. Dôležité je, že sa preukázalo, že NDP52 sa ko-lokalizuje s TAU v myších neurónoch a priama interakcia medzi fosfo-TAU a NDP52 bola preukázaná koimunoprecipitačnými experimentmi u myší aj vzoriek AD, čo poukazuje na jeho úlohu pri degradácii TAU. Je zaujímavé, že umlčanie NDP52, p62 alebo NRF2 v neurónoch viedlo k zvýšeniu fosfo-TAU [113], [118]. Navyše, zvýšené intraneuronálne APP agregáty boli nájdené v hipokampe APP/PS1AE9 myší, keď NRF2 chýbal. To korelovalo so zmenenými autofagickými markermi, vrátane zvýšených pomerov fosfo-mTOR/mTOR a fosfo-p70S6k/p70S6k (indikujúcich inhibíciu autofágie), zvýšenými hladinami prekatepsínu D a väčším počtom multivezikulárnych teliesok [119]. U myší koexprimujúcich ľudský APP (V717I) a TAU (P301L) nedostatok NRF2 viedol k zvýšeným hladinám celkového a fosfo-TAU v nerozpustnej frakcii a zvýšeným intraneuronálnym APP agregátom spolu so zníženými neurónovými hladinami p62, NDP52, ULK1, ATG5 a GABARAPL1. Spoločná lokalizácia medzi adaptorovým proteínom p62 a APP alebo TAU bola znížená v neprítomnosti NRF2 [36]. Celkovo tieto výsledky zdôrazňujú dôležitosť NRF2 v neurónovej autofágii.
Rôzne transkripčné faktory pôsobia koordinovane na moduláciu proteostázy
V podmienkach ustáleného stavu je proteostáza riadená prostredníctvom interakcií proteín-proteín a posttranslačných modifikácií, čím sa dosiahne rýchla odpoveď. Bunková adaptácia však vyžaduje transkripčnú reguláciu génov UPR, UPS a autofágie. Vzhľadom na to, že nervové bunky sú nepretržite vystavené toxickým útokom nízkeho stupňa, vrátane oxidačného a proteotoxického stresu, posilnenie proteostázy indukovanej transkripčnou moduláciou môže pomôcť zabrániť degenerácii mozgu.
V prípade UPR bude mať aktivácia každého z troch ramien nakoniec za následok transkripčnú indukciu určitých génov (prehľad v [43]). Napríklad fragment odvodený od ATF6 (ATF6f) sa viaže na prvky ER-stresovej odpovede (ERSE) a indukuje expresiu niekoľkých génov, vrátane XBPI, BIP a CHOP. Okrem toho signalizácia PERK vedie k aktivácii transkripčného faktora ATF4, ktorý riadi expresiu viacerých génov súvisiacich s UPR a niektorých ďalších vrátane cieľových génov NRF2 Hmox1 a p62. Nakoniec, aktivácia IRE1 vedie k vytvoreniu aktívneho transkripčného faktora, zostrihnutého XBP1 (XBP1s), ktorý riadi transkripciu génov kódujúcich proteíny zapojené do skladania proteínov.
Na druhej strane sa ukázalo, že NRF1 je nevyhnutný pre expresiu proteazomálneho génu v mozgu, pretože myši s knockoutom Nrf1 vykazovali zníženú expresiu génov kódujúcich rôzne podjednotky jadra 20S, ako aj regulačný komplex 19S spolu s poruchou proteazomálnej funkcie [90 ]. NRF1 aj NRF2 sa viažu na sekvencie ARE v promótorových oblastiach svojich cieľových génov, čo naznačuje, že majú prekrývajúce sa transkripčné aktivity, hoci sa líšia vo svojich regulačných mechanizmoch a bunkovej lokalizácii [120].
Transkripčné faktory rodiny Forkhead box O (FOXO) riadia expresiu viacerých génov súvisiacich s autofágiou. Podobne ako v prípade NRF2, existuje viacero vrstiev regulácie aktivity členov FOXO, ktoré môžu byť vyvolané nutričným alebo oxidačným stresom [121]. Nakoniec, transkripčný faktor TFEB, považovaný za hlavný regulátor lyzozomálnej biogenézy, hrá kľúčovú úlohu pri regulácii autofágie v podmienkach nutričného stresu. Inhibícia mTORC1 teda vedie k jadrovej translokácii TFEB a indukcii expresie autofágových génov [122].
Celkovo existencia rôznych transkripčných regulátorov týchto strojov tiež naznačuje presluchy a čiastočne redundantné mechanizmy, ktoré môžu zabezpečiť proteostázu za rôznych okolností. V súlade s tým môže mať NRF2 relevantnú úlohu v tkanivách, ktoré podporujú vysoké úrovne oxidačného stresu. Napríklad NRF2 vyvolaný oxidačným stresom môže fungovať v podmienkach bohatých na živiny, aby transkripčne upreguloval autofágiu, podobne ako to, čo sa zistilo pre TFEB v podmienkach hladovania. Okrem toho mozog funguje do značnej miery v podmienkach bohatých na živiny, čo predstavuje NRF2 ako relevantný mechanizmus na aktiváciu autofágie v neurónoch.
Sľubný �terapeutický potenciál pre NRF2 pri proteinopatiách
V posledných rokoch sa dosiahol veľký pokrok v znalostiach regulačných úloh UPR, UPS a autofágie na aktivitu NRF2, ako aj recipročnej transkripcii komponentov týchto troch systémov sprostredkovanej NRF2. Preto môžu vzniknúť nové terapeutické možnosti založené na využití NRF2 ako kľúčového regulátora klírensu proteínov pri neurodegeneratívnych ochoreniach.
Kľúčovou zostávajúcou otázkou však je, či bude užitočné alebo škodlivé zvýšiť hladiny NRF2 v mozgu. Analýza epidemiologických údajov môže poskytnúť čiastočnú odpoveď, pretože naznačuje, že gén NFE2L2 je vysoko polymorfný a niektoré jednonukleotidové polymorfizmy nachádzajúce sa v jeho promótorovej regulačnej oblasti môžu poskytnúť celý rad �fyziologických� variabilit v génovej expresii na úrovni populácie a niektorých haplotypov. boli spojené so zníženým rizikom a/alebo oneskoreným nástupom AD, PD alebo ALS [123]. Okrem toho, ako diskutovali Hayes a kolegovia [124], účinok NRF2 môže mať odpoveď v tvare písmena U, čo znamená, že príliš nízke hladiny NRF2 môžu viesť k strate cytoprotekcie a zvýšenej náchylnosti na stresory, zatiaľ čo príliš veľa NRF2 môže narušiť homeostatickú rovnováhu smerom k redukčný scenár (redukčný stres), ktorý by uprednostňoval nesprávne poskladanie a agregáciu proteínov. Nízke hladiny NRF2 v mozgu podporujú myšlienku, že mierna upregulácia môže byť dostatočná na dosiahnutie prínosu za patologických podmienok. V skutočnosti bola ochranná úloha farmakologickej aktivácie klírensu proteínu sprostredkovaná NRF2 preukázaná v rôznych neurodegeneračných bunkových kultúrach a in vivo modeloch.
SFN je farmakologický aktivátor NRF2, o ktorom sa preukázalo, že indukuje expresiu proteazomálneho a autofágového génu [95], [36]. Je zaujímavé, že Jo a kolegovia preukázali, že SFN znižuje hladiny fosforylovaného TAU a zvyšuje Beclin-1 a LC3-II, čo naznačuje, že aktivácia NRF2 môže uľahčiť degradáciu tohto toxického proteínu prostredníctvom autofágie [113]. Okrem toho bola degradácia mHtt zvýšená pomocou SFN, a to bolo zvrátené použitím MG132, čo naznačuje proteazomálnu degradáciu tohto toxického proteínu [95]. Autofágia sprostredkovaná degradácia fosfo- a nerozpustného-TAU bola hlásená s organickým flavonoidom fisetínom. Táto zlúčenina bola schopná vyvolať autofágiu súčasnou podporou aktivácie a jadrovej translokácie TFEB aj NRF2 spolu s niektorými z jej cieľových génov. Tejto reakcii zabránilo umlčanie TFEB alebo NRF2 [125]. Bott a kolegovia uviedli priaznivé účinky simultánneho aktivátora NRF2, NRF1 a HSF1 na proteínovú toxicitu pri spinálnej a bulbárnej svalovej atrofii, neurodegeneratívnej poruche spôsobenej expanziou CAG opakovaní kódujúcich polyglutamín, v ktorých sú prítomné proteínové agregáty [126]. Potenciál aktivácie NRF2 na liečbu neurodegeneratívnych porúch bol preukázaný schválením BG-12, perorálnej formulácie dimetylfumarátu induktora NRF2 (DMF), na liečbu roztrúsenej sklerózy [127], [128]. Úspech DMF pri autoimunitných ochoreniach so silnou zápalovou zložkou naznačuje, že neurodegeneratívne ochorenia môžu mať prospech z premiestnenia tohto lieku. V nedávnej predklinickej štúdii modelu y-synukleinopatie PD sa ukázalo, že DMF je neuroprotektívny, čiastočne v dôsledku jeho indukcie autofágie [129]. Štúdie uvádzajúce priaznivé účinky NRF2 na neurodegeneráciu, ale nezameriavajúce sa na jeho účinok na klírens proteínov, sú ešte hojnejšie (komplexný prehľad nájdete v [7]). To je celkom relevantné, pretože to poukazuje na viaceré škodlivé procesy, ktoré môžu byť súčasne zacielené jediným zásahom v NRF2, vrátane oxidačného stresu, neurozápalu alebo mitochondriálnej dysfunkcie. Budúca práca však bude potrebná na definitívne určenie, či farmakologická aktivácia NRF2 môže byť platnou stratégiou na uľahčenie degradácie toxických proteínov v mozgu.
Ako už bolo vysvetlené, zhoršený GSK-3? aktivita bola hlásená pri neurodegeneratívnych ochoreniach a špekulovalo sa, že následné zníženie NRF2 môže byť čiastočne zodpovedné za škodlivý výsledok. Za týchto patologických podmienok by inhibítory GSK-3 mohli tiež spolupracovať na zvýšení hladín NRF2 a proteostázy. Priaznivé účinky inhibítorov GSK-3 boli zaznamenané v rôznych modeloch neurodegenerácie a čo je zaujímavejšie, represia GSK-3 dokázala znížiť hladiny toxických proteínov [130], [131], [132], [133]. Aj keď sa zatiaľ nepozorovali žiadne priame spojenia medzi inhibíciou GSK-3 a transkripčnou reguláciou génov podporujúcich proteostázu NRF2, je rozumné špekulovať, že zníženie aktivity GSK-3 by viedlo k zvýšeniu hladín NRF2, čo by nakoniec viedlo k posilneniu proteostáza.
Transkripčná aktivita NRF2, ako aj bunková kapacita udržiavať proteostázu klesá s vekom, čo je hlavný rizikový faktor pre rozvoj neurodegeneratívnych ochorení. Je rozumné si myslieť, že posilnenie NRF2 a následne proteostáza by prinajmenšom oddialilo akumuláciu proteínových agregátov a neurodegeneráciu. Ošetrenie ľudských senescentných fibroblastov triterpenoidom kyseliny 18a-glycyrrhetínovej (18p-GA) skutočne podporovalo aktiváciu NRF2, čo viedlo k indukcii proteazómu a predĺženiu životnosti. Táto štúdia naznačuje, že farmakologická aktivácia NRF2 je možná aj v neskoršom veku [86]. Okrem toho neskoršia štúdia ukázala, že táto zlúčenina sprostredkovala aktiváciu SKN-1 a proteazómu v C. elegans s priaznivými účinkami na progresiu AD v relevantných modeloch nematód [134].
Po zvážení všetkých vecí sa zdá, že indukcia génov súvisiacich s proteostázou sprostredkovaná NRF2 je prospešná pri rôznych proteinopatiách.
Sulforaphane a jeho účinky na rakovinu, úmrtnosť, starnutie, mozog a správanie, srdcové choroby a ďalšie
Izotiokyanáty sú niektoré z najdôležitejších rastlinných zlúčenín, ktoré môžete získať vo svojej strave. V tomto videu pre nich robím najkomplexnejší prípad, aký kedy bol vyrobený. Krátka doba pozornosti? Preskočte na svoju obľúbenú tému kliknutím na jeden z časových bodov nižšie. Úplná časová os nižšie.
Kľúčové sekcie:
00:01:14 – Rakovina a úmrtnosť
00:19:04 – Starnutie
00:26:30 – Mozog a správanie
00:38:06 – Záverečná rekapitulácia
00:40:27 – Dávka
Úplná časová os:
00:00:34 – Predstavenie sulforafanu, hlavné zameranie videa.
00:01:14 – Spotreba krížovej zeleniny a zníženie úmrtnosti zo všetkých príčin.
00:02:12 – Riziko rakoviny prostaty.
00:02:23 – Riziko rakoviny močového mechúra.
00:02:34 – Riziko rakoviny pľúc u fajčiarov.
00:02:48 – Riziko rakoviny prsníka.
00:03:13 – Hypotetické: čo ak už máte rakovinu? (intervenčné)
00:03:35 – Pravdepodobný mechanizmus, ktorý riadi asociatívne údaje o rakovine a úmrtnosti.
00:04:38 – Sulforafán a rakovina.
00:05:32 – Dôkazy na zvieratách ukazujúce silný účinok extraktu z výhonkov brokolice na vývoj nádoru močového mechúra u potkanov.
00:06:06 – Účinok priamej suplementácie sulforafanu u pacientov s rakovinou prostaty.
00:07:09 – Bioakumulácia izotiokyanátových metabolitov v aktuálnom prsnom tkanive.
00:08:32 – Inhibícia kmeňových buniek rakoviny prsníka.
00:08:53 – Lekcia dejepisu: Brassica bola preukázaná ako zdraviu prospešné už v starovekom Ríme.
00:09:16 – Schopnosť sulforafanu zvyšovať vylučovanie karcinogénov (benzén, akroleín).
00:09:51 – NRF2 ako genetický prepínač prostredníctvom prvkov antioxidačnej odozvy.
00:10:10 – Ako aktivácia NRF2 zvyšuje vylučovanie karcinogénu cez glutatión-S-konjugáty.
00:10:34 – Ružičkový kel zvyšuje glutatión-S-transferázu a znižuje poškodenie DNA.
00:11:20 – Nápoj z brokolicových klíčkov zvyšuje vylučovanie benzénu o 61 %.
00:13:31 – Homogenát brokolicových klíčkov zvyšuje antioxidačné enzýmy v horných dýchacích cestách.
00:15:45 – Spotreba krížovej zeleniny a úmrtnosť na srdcové choroby.
00:16:55 – Prášok z brokolicových klíčkov zlepšuje krvné lipidy a celkové riziko srdcových chorôb u diabetikov 2. typu.
00:19:04 – Začiatok úseku starnutia.
00:19:21 – Diéta obohatená o sulforafan zvyšuje životnosť chrobákov z 15 na 30 % (za určitých podmienok).
00:20:34 – Význam nízkeho zápalu pre dlhovekosť.
00:22:05 – Zdá sa, že krížová zelenina a prášok z brokolicových klíčkov znižujú množstvo zápalových markerov u ľudí.
00:23:40 – Rekapitulácia v polovici videa: rakovina, starnutie
00:24:14 – Štúdie na myšiach naznačujú, že sulforafan môže zlepšiť adaptívnu imunitnú funkciu v starobe.
00:25:18 – Sulforaphane zlepšil rast vlasov u myšieho modelu plešatenia. Snímka o 00:26:10.
00:26:30 – Začiatok sekcie mozog a správanie.
00:27:18 – Vplyv extraktu z brokolicových klíčkov na autizmus.
00:27:48 – Účinok glukorafanínu na schizofréniu.
00:28:17 – Začiatok diskusie o depresii (pravdepodobný mechanizmus a štúdie).
00:31:21 – Štúdia na myšiach s použitím 10 rôznych modelov depresie vyvolanej stresom ukazuje, že sulforafan je podobne účinný ako fluoxetín (prozac).
00:32:00 – Štúdia ukazuje, že priame požitie glukorafanínu u myší je podobne účinné pri prevencii depresie zo sociálneho modelu stresu.
00:33:01 – Začiatok sekcie neurodegenerácie.
00:33:30 – Sulforafán a Alzheimerova choroba.
00:33:44 – Sulforaphane a Parkinsonova choroba.
00:33:51 - Sulforaphane a Hungtingtonova choroba.
00:34:13 – Sulforaphane zvyšuje proteíny tepelného šoku.
00:34:43 – Začiatok sekcie traumatického poranenia mozgu.
00:35:01 – Sulforaphane podaný ihneď po TBI zlepšuje pamäť (štúdia na myšiach).
00:35:55 – Sulforafán a plasticita neurónov.
00:36:32 – Sulforaphane zlepšuje učenie v modeli diabetu typu II u myší.
00:37:19 – Sulforaphane a Duchennova svalová dystrofia.
00:37:44 – Inhibícia myostatínu vo svalových satelitných bunkách (in vitro).
00:38:06 – Neskoré video rekapitulácia: úmrtnosť a rakovina, poškodenie DNA, oxidačný stres a zápal, vylučovanie benzénu, kardiovaskulárne ochorenia, diabetes typu II, účinky na mozog (depresia, autizmus, schizofrénia, neurodegenerácia), dráha NRF2.
00:40:27 – Myšlienky na to, ako zistiť dávku brokolicových klíčkov alebo sulforafanu.
00:41:01 – Anekdoty o klíčení doma.
00:43:14 – O teplotách varenia a aktivite sulforafanu.
00:43:45 – Premena sulforafanu z glukorafanínu črevnými baktériami.
00:44:24 – Doplnky fungujú lepšie v kombinácii s aktívnou myrozinázou zo zeleniny.
00:44:56 – Techniky varenia a krížová zelenina.
00:46:06 – Izotiokyanáty ako struma.
Faktor 2 súvisiaci s erytroidným faktorom 2 (NF-E2), inak známy ako Nrf2, je transkripčný faktor, ktorý reguluje expresiu rôznych antioxidačných a detoxikačných enzýmov. Výskumné štúdie tiež preukázali jeho úlohu pri kontrole oxidačného stresu. Väčšina neurodegeneratívnych ochorení, ako je Alzheimerova choroba a Parkinsonova choroba, sa vyznačuje oxidačným stresom a chronickým zápalom, čo sú spoločné ciele Prístupy k liečbe Nrf2. Dr. Alex Jimenez DC, CCST Insight
Záverečné poznámky
Transkripčný faktor NRF2 riadi proteostatickú odpoveď snímaním a moduláciou zmien v UPR, UPS a autofágii (obr. 4). V dôsledku toho sa ukázalo, že nedostatok NRF2 zhoršuje proteinopatiu, čo naznačuje, že NRF2 je nevyhnutný pre optimálny klírens proteínov. Všetci spolu môžeme špekulovať, že NRF2 môže byť zaujímavým terapeutickým cieľom pre proteinopatie.
Obrázok 4 NRF2 ako rozbočovač spájajúci núdzové signály odvodené od proteotoxických látok s ochrannou transkripčnou odpoveďou. Akumulácia neposkladaných/chybne poskladaných proteínov povedie k aktivácii reakcie na rozbalený proteín (UPR) v ER. Aktivácia PERK alebo MAPK môže viesť k transkripčnej indukcii ER-rezidentného Gpx8 a niekoľkých enzýmov regulujúcich hladiny GSH, čo je kritické na zabezpečenie správneho skladania proteínov. Proteínové agregáty inhibujú aktivitu proteazómu (UPS), čím sa pravdepodobne vyhýbajú degradácii NRF2. Ukázalo sa, že NRF2 špecificky moduluje transkripciu génov Psma3, Psma6, Psmb1, Psmb5 a Pomp. Niekoľko ďalších podjednotiek bolo upregulovaných spôsobom závislým od NRF2 v reakcii na D3T, čím sa pravdepodobne rozšíril zoznam proteazómových podjednotiek regulovaných NRF2. Autofágia je hlavnou cestou degradácie proteínových agregátov. Autofágia tiež reguluje NRF2 a spája túto degradačnú dráhu s transkripčnou indukciou NRF2 p62, Ndp52, Ulk1, Atg2b, Atg4c, Atg5, Atg7 a Gabarapl1.
Podľa vyššie uvedeného článku, zatiaľ čo symptómy neurodegeneratívnych ochorení možno liečiť rôznymi spôsobmi liečby, výskumné štúdie preukázali, že aktivácia Nrf2 môže byť užitočným liečebným prístupom. Pretože Aktivátory Nrf2 sa zameriavajú na široké mechanizmy ochoreniaVšetky neurodegeneratívne ochorenia môžu mať prospech z použitia transkripčného faktora Nrf2. Zistenia Nrf2 spôsobili revolúciu v liečbe neurodegeneratívnych ochorení. Rozsah našich informácií je obmedzený na chiropraktické a zdravotné problémy chrbtice. Ak chcete prediskutovať túto tému, neváhajte sa opýtať Dr. Jimeneza alebo nás kontaktujte na adrese�915-850-0900 .
Ďalšia téma diskusie: Zmiernenie bolesti kolena bez operácie
Bolesť kolena je dobre známym príznakom, ktorý sa môže vyskytnúť v dôsledku rôznych zranení a/alebo stavov kolena, vrátane�športové zranenia. Koleno je jedným z najzložitejších kĺbov v ľudskom tele, pretože je tvorené priesečníkom štyroch kostí, štyroch väzov, rôznych šliach, dvoch meniskov a chrupavky. Podľa Americkej akadémie rodinných lekárov medzi najčastejšie príčiny bolesti kolena patrí patelárna subluxácia, patelárna tendinitída alebo skokanovo koleno a Osgood-Schlatterova choroba. Hoci bolesť kolena sa s najväčšou pravdepodobnosťou vyskytuje u ľudí starších ako 60 rokov, bolesť kolena sa môže vyskytnúť aj u detí a dospievajúcich. Bolesť kolena sa môže liečiť doma podľa metód RICE, avšak vážne poranenia kolena môžu vyžadovať okamžitú lekársku pomoc vrátane chiropraktickej starostlivosti. �
Oxidačný stres je opísaný ako poškodenie buniek spôsobené voľnými radikálmi alebo nestabilnými molekulami, ktoré môžu v konečnom dôsledku ovplyvniť zdravú funkciu. Ľudské telo vytvára voľné radikály na neutralizáciu baktérií a vírusov, avšak vonkajšie faktory, ako je kyslík, znečistenie a žiarenie, môžu často tiež produkovať voľné radikály. Oxidačný stres je spojený s mnohými zdravotnými problémami.
Oxidačný stres a iné stresory zapínajú vnútorné ochranné mechanizmy, ktoré môžu pomôcť regulovať antioxidačnú reakciu ľudského tela. Nrf2 je proteín, ktorý sníma úrovne oxidačného stresu a umožňuje bunkám chrániť sa pred vnútornými a vonkajšími faktormi. Ukázalo sa tiež, že Nrf2 pomáha regulovať gény zapojené do produkcie antioxidačných enzýmov a génov odozvy na stres. Účelom nižšie uvedeného článku je vysvetliť účinky Nrf2 pri rakovine.
abstraktné
Dráha Keap1-Nrf2 je hlavným regulátorom cytoprotektívnych reakcií na oxidačný a elektrofilný stres. Hoci bunkové signálne dráhy spúšťané transkripčným faktorom Nrf2 zabraňujú iniciácii a progresii rakoviny v normálnych a premalígnych tkanivách, v úplne malígnych bunkách aktivita Nrf2 poskytuje rastovú výhodu zvýšením chemorezistencie rakoviny a zvýšením rastu nádorových buniek. V tomto grafickom prehľade poskytujeme prehľad o dráhe Keap1-Nrf2 a jej dysregulácii v rakovinových bunkách. Stručne tiež zhrnieme dôsledky konštitutívnej aktivácie Nrf2 v rakovinových bunkách a ako to možno využiť v génovej terapii rakoviny.
Kľúčové slová:Nrf2, Keap1, rakovina, prvok antioxidačnej odozvy, génová terapia
úvod
Dráha Keap1-Nrf2 je hlavným regulátorom cytoprotektívnych reakcií na endogénne a exogénne stresy spôsobené reaktívnymi formami kyslíka (ROS) a elektrofilmi [1]. Kľúčovými signálnymi proteínmi v dráhe sú transkripčný faktor Nrf2 (faktor 2 súvisiaci s erytroidným faktorom 2), ktorý sa viaže spolu s malými proteínmi Maf na prvok antioxidačnej odozvy (ARE) v regulačných oblastiach cieľových génov, a Keap1 (Kelch ECH asociačný proteín 1), represorový proteín, ktorý sa viaže na Nrf2 a podporuje jeho degradáciu prostredníctvom ubikvitínovej proteazómovej dráhy (obr. 1). Keap1 je proteín veľmi bohatý na cysteín, myšací Keap1 má celkovo 25 a ľudský 27 cysteínových zvyškov, z ktorých väčšina môže byť modifikovaná in vitro rôznymi oxidantmi a elektrofilmi [2]. Ukázalo sa, že tri z týchto zvyškov, C151, C273 a C288, hrajú funkčnú úlohu zmenou konformácie Keap1, čo vedie k jadrovej translokácii Nrf2 a následnej expresii cieľového génu [3] (obr. 1). Presný mechanizmus, ktorým cysteínové modifikácie v Keap1 vedú k aktivácii Nrf2, nie je známy, ale dva prevládajúce, ale vzájomne sa nevylučujúce modely sú (1) model „závesu a západky“, v ktorom sú modifikácie Keap1 v tiolových zvyškoch nachádzajúcich sa v IVR Keap1 narúšajú interakciu s Nrf2, čo spôsobuje nesprávne zarovnanie lyzínových zvyškov v Nrf2, ktoré už nemôžu byť polyubikvitinylované, a (2) model, v ktorom modifikácia tiolu spôsobuje disociáciu Cul3 od Keap1 [3]. V oboch modeloch je induktorom modifikovaný a na Nrf2 viazaný Keap1 inaktivovaný a následne novosyntetizované proteíny Nrf2 obchádzajú Keap1 a translokujú sa do jadra, viažu sa na ARE a riadia expresiu cieľových génov Nrf2, ako je NAD(P)H chinónoxidoreduktáza 1 (NQO1), hemoxygenáza 1 (HMOX1), glutamát-cysteín ligáza (GCL) a glutatión S transferázy (GST) (obr. 2). Okrem modifikácií tiolov Keap1, ktoré vedú k indukcii cieľového génu Nrf2, sa proteíny ako p21 a p62 môžu viazať na Nrf2 alebo Keap1, čím narúšajú interakciu medzi Nrf2 a Keap1 [1], [3] (obr. 3).
Obr. 1. Štruktúry Nrf2 a Keap1 a cysteínový kód. (A) Nrf2 pozostáva z 589 aminokyselín a má šesť evolučne vysoko konzervovaných domén, Neh1-6. Neh1 obsahuje motív bZip, štruktúru základnej oblasti � leucínového zipsu (L-Zip), kde základná oblasť je zodpovedná za rozpoznávanie DNA a L-Zip sprostredkúva dimerizáciu s malými proteínmi Maf. Neh6 funguje ako degron na sprostredkovanie degradácie Nrf2 v jadre. Neh4 a 5 sú transaktivačné domény. Neh2 obsahuje motívy ETGE a DLG, ktoré sú potrebné na interakciu s Keap1, a hydrofilnú oblasť lyzínových zvyškov (7 K), ktoré sú nevyhnutné pre polyubikvitináciu závislú od Keap1 a degradáciu Nrf2. (B) Keap1 pozostáva zo 624 aminokyselinových zvyškov a má päť domén. Dva motívy interakcie proteín-proteín, doména BTB a doména Kelch, sú oddelené intervenujúcou oblasťou (IVR). BTB doména spolu s N-koncovou časťou IVR sprostredkúva homodimerizáciu Keap1 a väzbu s Cullin3 (Cul3). Kelchova doména a C-koncová oblasť sprostredkúvajú interakciu s Neh2. (C) Nrf2 interaguje s dvoma molekulami Keap1 prostredníctvom svojich motívov Neh2 ETGE a DLG. ETGE aj DLG sa viažu na podobné miesta na spodnom povrchu motívu Keap1 Kelch. (D) Keap1 je bohatý na cysteínové zvyšky s 27 cysteínmi v ľudskom proteíne. Niektoré z týchto cysteínov sa nachádzajú v blízkosti bázických zvyškov, a preto sú vynikajúcimi cieľmi elektrofilov a oxidantov. Modifikačný vzor cysteínových zvyškov elektrofilmi je známy ako cysteínový kód. Hypotéza cysteínového kódu navrhuje, že štrukturálne odlišné činidlá aktivujúce Nrf2 ovplyvňujú rôzne cysteíny Keap1. Cysteínové modifikácie vedú ku konformačným zmenám v Keap1, ktoré narúšajú interakciu medzi doménami Nrf2 DLG a Keap1 Kelch, čím inhibujú polyubikvitináciu Nrf2. Funkčná dôležitosť Cys151, Cys273 a Cys288 bola preukázaná, pretože Cys273 a Cys288 sú potrebné na potlačenie Nrf2 a Cys151 na aktiváciu Nrf2 induktormi [1], [3].
Obr. 2. Signálna dráha Nrf2-Keap1. (A a B) v bazálnych podmienkach sa dve molekuly Keap1 viažu na Nrf2 a Nrf2 je polyubikvitylovaný komplexom E3 ligázy na báze Cul3. Táto polyubikvitilácia vedie k rýchlej degradácii Nrf2 proteazómom. Malá časť Nrf2 uniká z inhibičného komplexu a hromadí sa v jadre, aby sprostredkovala bazálnu génovú expresiu závislú od ARE, čím sa udržiava bunková homeostáza. (C) Za stresových podmienok induktory modifikujú cysteíny Keap1, čo vedie k inhibícii ubikvitylácie Nrf2 prostredníctvom disociácie inhibičného komplexu. (D) Podľa modelu pántu a západky vedie modifikácia špecifických cysteínových zvyškov Keap1 ku konformačným zmenám v Keap1, čo vedie k oddeleniu motívu Nrf2 DLG od Keap1. Ubikvitinácia Nrf2 je narušená, ale väzba s motívom ETGE zostáva. (E) V modeli disociácie Keap1-Cul3 je väzba Keap1 a Cul3 prerušená v reakcii na elektrofily, čo vedie k úniku Nrf2 z ubikvitinačného systému. V oboch navrhovaných modeloch je induktorom modifikovaný a na Nrf2 viazaný Keap1 inaktivovaný a následne novosyntetizované proteíny Nrf2 obchádzajú Keap1 a premiestňujú sa do jadra, viažu sa na prvok Antioxidačnej odozvy (ARE) a riadia expresiu cieľa Nrf2. gény ako NQO1, HMOX1, GCL a GST [1], [3].
Obr. 3. Mechanizmy konštitutívnej jadrovej akumulácie Nrf2 pri rakovine. (A) Somatické mutácie v Nrf2 alebo Keap1 narúšajú interakciu týchto dvoch proteínov. V Nrf2 mutácie ovplyvňujú motívy ETGE a DLG, ale v Keap1 sú mutácie distribuované rovnomernejšie. Okrem toho aktivácia onkogénu, ako je KrasG12D[5], alebo narušenie supresorov nádorov, ako je PTEN [11], môže viesť k transkripčnej indukcii Nrf2 a zvýšeniu jadrového Nrf2. (B) Hypermetylácia promótora Keap1 pri rakovine pľúc a prostaty vedie k zníženiu expresie Keap1 mRNA, čo zvyšuje jadrovú akumuláciu Nrf2 [6], [7]. (C) Pri familiárnom papilárnom obličkovom karcinóme vedie strata aktivity enzýmu fumaráthydratázy k akumulácii fumarátu a ďalej k sukcinácii cysteínových zvyškov Keap1 (2SC). Táto posttranslačná modifikácia vedie k narušeniu interakcie Keap1-Nrf2 a jadrovej akumulácii Nrf2 [8], [9]. (D) Akumulácia disruptorových proteínov, ako sú p62 a p21, môže narušiť väzbu Nrf2-Keap1 a vedie k zvýšeniu jadrového Nrf2. p62 sa viaže na Keap1 a prekrýva väzobné vrecko pre Nrf2 a p21 priamo interaguje s motívmi DLG a ETGE Nrf2, čím konkuruje Keap1 [10].
Mechanizmy aktivácie a dysregulácie Nrf2 pri rakovine
Hoci cytoprotekcia poskytovaná aktiváciou Nrf2 je dôležitá pre chemoprevenciu rakoviny v normálnych a premalígnych tkanivách, v úplne malígnych bunkách aktivita Nrf2 poskytuje rastovú výhodu zvýšením chemorezistencie rakoviny a zvýšením rastu nádorových buniek [4]. Bolo opísaných niekoľko mechanizmov, ktorými je signálna dráha Nrf2 konštitutívne aktivovaná pri rôznych rakovinách: (1) somatické mutácie v Keap1 alebo väzbovej doméne Keap1 Nrf2 narušujúce ich interakciu; (2) epigenetické umlčanie expresie Keap1 vedúce k defektnej represii Nrf2; (3) akumulácia disruptorových proteínov, ako je p62, čo vedie k disociácii komplexu Keap1-Nrf2; (4) transkripčná indukcia Nrf2 onkogénnymi K-Ras, B-Raf a c-Myc; a (5) posttranslačnú modifikáciu Keap1 cysteínov sukcinyláciou, ktorá sa vyskytuje pri familiárnom papilárnom renálnom karcinóme v dôsledku straty enzýmovej aktivity fumarát hydratázy [3], [4], [5], [6], [7], [ 8], [9], [10] (obr. 3). Konštitutívne hojný proteín Nrf2 spôsobuje zvýšenú expresiu génov zapojených do metabolizmu liečiv, čím zvyšuje odolnosť voči chemoterapeutickým liečivám a rádioterapii. Okrem toho vysoká hladina proteínu Nrf2 je spojená so zlou prognózou rakoviny [4]. Nadmerne aktívny Nrf2 tiež ovplyvňuje bunkovú proliferáciu nasmerovaním glukózy a glutamínu smerom k anabolickým dráham, ktoré zvyšujú syntézu purínov a ovplyvňujú dráhu pentózofosfátu na podporu bunkovej proliferácie [11] (obr. 4).
Obr. 4. Dvojitá úloha Nrf2 pri tumorigenéze. Za fyziologických podmienok postačujú nízke hladiny jadrového Nrf2 na udržanie bunkovej homeostázy. Nrf2 inhibuje iniciáciu nádoru a rakovinové metastázy elimináciou karcinogénov, ROS a iných látok poškodzujúcich DNA. Počas tumorigenézy vedie akumulujúce sa poškodenie DNA ku konštitutívnej hyperaktivite Nrf2, ktorá pomáha autonómnym malígnym bunkám vydržať vysoké hladiny endogénneho ROS a vyhnúť sa apoptóze. Trvalo zvýšené hladiny jadrového Nrf2 aktivujú okrem cytoprotektívnych génov aj metabolické gény, ktoré prispievajú k metabolickému preprogramovaniu a zvýšeniu bunkovej proliferácie. Rakoviny s vysokými hladinami Nrf2 sú spojené so zlou prognózou v dôsledku rádiovej a chemorezistencie a agresívnej proliferácie rakovinových buniek. Aktivita dráhy Nrf2 je teda ochranná v skorých štádiách tumorigenézy, ale škodlivá v neskorších štádiách. Preto na prevenciu rakoviny zostáva zvýšenie aktivity Nrf2 dôležitým prístupom, zatiaľ čo na liečbu rakoviny je žiaduca inhibícia Nrf2 [4], [11].
Vzhľadom na to, že vysoká aktivita Nrf2 sa bežne vyskytuje v rakovinových bunkách s nepriaznivými výsledkami, existuje potreba terapií na inhibíciu Nrf2. Bohužiaľ, v dôsledku štrukturálnej podobnosti s niektorými ďalšími členmi rodiny bZip je vývoj špecifických inhibítorov Nrf2 náročnou úlohou a doteraz bolo publikovaných len niekoľko štúdií inhibície Nrf2. Skríningom prírodných produktov Ren a spol. [12] identifikovali antineoplastickú zlúčeninu brusatol ako inhibítor Nrf2, ktorý zvyšuje chemoterapeutickú účinnosť cisplatiny. Okrem toho sa na inhibíciu Nrf3 v rakovinových bunkách použili inhibítory PI11K [13], [2] a Nrf14 siRNA [2]. Nedávno sme použili alternatívny prístup, známy ako génová terapia samovraždy rakoviny, na zacielenie rakovinových buniek s vysokými hladinami Nrf2. Lentivírusové vektory riadené Nrf2 [15] obsahujúce tymidínkinázu (TK) sa prenesú do rakovinových buniek s vysokou aktivitou ARE a bunky sa ošetria proliečivom, ganciklovirom (GCV). GCV sa metabolizuje na GCV-monofosfát, ktorý je ďalej fosforylovaný bunkovými kinázami na toxickú trifosfátovú formu [16] (obr. 5). To vedie k účinnému zabíjaniu nielen nádorových buniek obsahujúcich TK, ale aj susedných buniek v dôsledku efektu okoloidúcich [17]. Génová terapia TK/GCV regulovaná ARE môže byť ďalej posilnená kombináciou protirakovinovej chemoterapeutickej látky doxorubicínu s liečbou [16], čo podporuje názor, že tento prístup by mohol byť užitočný v spojení s tradičnými terapiami.
Obr. 5. Samovražedná génová terapia. Konštitutívna jadrová akumulácia Nrf2 v rakovinových bunkách sa môže využiť pomocou vírusového vektora riadeného Nrf2 na génovú terapiu samovraždy rakoviny [16]. V tomto prístupe sa lentivírusový vektor (LV) exprimujúci tymidínkinázu (TK) pod minimálnym promótorom SV40 so štyrmi ARE transdukuje do buniek adenokarcinómu pľúc. Vysoké hladiny jadrového Nrf2 vedú k robustnej expresii TK prostredníctvom väzby Nrf2. Bunky sú potom ošetrené proliečivom, ganciklovirom (GCV), ktorý je fosforylovaný TK. Trifosforylovaný GCV narúša syntézu DNA a vedie k účinnému zabíjaniu nielen nádorových buniek obsahujúcich TK, ale aj susedných buniek v dôsledku vedľajšieho účinku.
Nrf2 je hlavný regulátor, ktorý spúšťa produkciu silných antioxidantov v ľudskom tele, ktoré pomáhajú eliminovať oxidačný stres. Prostredníctvom dráhy Nrf2 sa aktivujú aj rôzne antioxidačné enzýmy, ako je superoxiddismutáza alebo SOD, glutatión a kataláza. Okrem toho niektoré fytochemikálie, ako je kurkuma, ashwagandha, bacopa, zelený čaj a ostropestrec, aktivujú Nrf2. Výskumné štúdie to zistili Aktivácia Nrf2 môže prirodzene zvýšiť ochranu buniek a obnoviť rovnováhu ľudského tela.
Dr. Alex Jimenez DC, CCST Insight
Sulforaphane a jeho účinky na rakovinu, úmrtnosť, starnutie, mozog a správanie, srdcové choroby a ďalšie
Izotiokyanáty sú niektoré z najdôležitejších rastlinných zlúčenín, ktoré môžete získať vo svojej strave. V tomto videu pre nich robím najkomplexnejší prípad, aký kedy bol vyrobený. Krátka doba pozornosti? Preskočte na svoju obľúbenú tému kliknutím na jeden z časových bodov nižšie. Úplná časová os nižšie.
Kľúčové sekcie:
00:01:14 – Rakovina a úmrtnosť
00:19:04 – Starnutie
00:26:30 – Mozog a správanie
00:38:06 – Záverečná rekapitulácia
00:40:27 – Dávka
Úplná časová os:
00:00:34 – Predstavenie sulforafanu, hlavné zameranie videa.
00:01:14 – Spotreba krížovej zeleniny a zníženie úmrtnosti zo všetkých príčin.
00:02:12 – Riziko rakoviny prostaty.
00:02:23 – Riziko rakoviny močového mechúra.
00:02:34 – Riziko rakoviny pľúc u fajčiarov.
00:02:48 – Riziko rakoviny prsníka.
00:03:13 – Hypotetické: čo ak už máte rakovinu? (intervenčné)
00:03:35 – Pravdepodobný mechanizmus, ktorý riadi asociatívne údaje o rakovine a úmrtnosti.
00:04:38 – Sulforafán a rakovina.
00:05:32 – Dôkazy na zvieratách ukazujúce silný účinok extraktu z výhonkov brokolice na vývoj nádoru močového mechúra u potkanov.
00:06:06 – Účinok priamej suplementácie sulforafanu u pacientov s rakovinou prostaty.
00:07:09 – Bioakumulácia izotiokyanátových metabolitov v aktuálnom prsnom tkanive.
00:08:32 – Inhibícia kmeňových buniek rakoviny prsníka.
00:08:53 – Lekcia dejepisu: Brassica bola preukázaná ako zdraviu prospešné už v starovekom Ríme.
00:09:16 – Schopnosť sulforafanu zvyšovať vylučovanie karcinogénov (benzén, akroleín).
00:09:51 – NRF2 ako genetický prepínač prostredníctvom prvkov antioxidačnej odozvy.
00:10:10 – Ako aktivácia NRF2 zvyšuje vylučovanie karcinogénu cez glutatión-S-konjugáty.
00:10:34 – Ružičkový kel zvyšuje glutatión-S-transferázu a znižuje poškodenie DNA.
00:11:20 – Nápoj z brokolicových klíčkov zvyšuje vylučovanie benzénu o 61 %.
00:13:31 – Homogenát brokolicových klíčkov zvyšuje antioxidačné enzýmy v horných dýchacích cestách.
00:15:45 – Spotreba krížovej zeleniny a úmrtnosť na srdcové choroby.
00:16:55 – Prášok z brokolicových klíčkov zlepšuje krvné lipidy a celkové riziko srdcových chorôb u diabetikov 2. typu.
00:19:04 – Začiatok úseku starnutia.
00:19:21 – Diéta obohatená o sulforafan zvyšuje životnosť chrobákov z 15 na 30 % (za určitých podmienok).
00:20:34 – Význam nízkeho zápalu pre dlhovekosť.
00:22:05 – Zdá sa, že krížová zelenina a prášok z brokolicových klíčkov znižujú množstvo zápalových markerov u ľudí.
00:23:40 – Rekapitulácia v polovici videa: rakovina, starnutie
00:24:14 – Štúdie na myšiach naznačujú, že sulforafan môže zlepšiť adaptívnu imunitnú funkciu v starobe.
00:25:18 – Sulforaphane zlepšil rast vlasov u myšieho modelu plešatenia. Snímka o 00:26:10.
00:26:30 – Začiatok sekcie mozog a správanie.
00:27:18 – Vplyv extraktu z brokolicových klíčkov na autizmus.
00:27:48 – Účinok glukorafanínu na schizofréniu.
00:28:17 – Začiatok diskusie o depresii (pravdepodobný mechanizmus a štúdie).
00:31:21 – Štúdia na myšiach s použitím 10 rôznych modelov depresie vyvolanej stresom ukazuje, že sulforafan je podobne účinný ako fluoxetín (prozac).
00:32:00 – Štúdia ukazuje, že priame požitie glukorafanínu u myší je podobne účinné pri prevencii depresie zo sociálneho modelu stresu.
00:33:01 – Začiatok sekcie neurodegenerácie.
00:33:30 – Sulforafán a Alzheimerova choroba.
00:33:44 – Sulforaphane a Parkinsonova choroba.
00:33:51 - Sulforaphane a Hungtingtonova choroba.
00:34:13 – Sulforaphane zvyšuje proteíny tepelného šoku.
00:34:43 – Začiatok sekcie traumatického poranenia mozgu.
00:35:01 – Sulforaphane podaný ihneď po TBI zlepšuje pamäť (štúdia na myšiach).
00:35:55 – Sulforafán a plasticita neurónov.
00:36:32 – Sulforaphane zlepšuje učenie v modeli diabetu typu II u myší.
00:37:19 – Sulforaphane a Duchennova svalová dystrofia.
00:37:44 – Inhibícia myostatínu vo svalových satelitných bunkách (in vitro).
00:38:06 – Neskoré video rekapitulácia: úmrtnosť a rakovina, poškodenie DNA, oxidačný stres a zápal, vylučovanie benzénu, kardiovaskulárne ochorenia, diabetes typu II, účinky na mozog (depresia, autizmus, schizofrénia, neurodegenerácia), dráha NRF2.
00:40:27 – Myšlienky na to, ako zistiť dávku brokolicových klíčkov alebo sulforafanu.
00:41:01 – Anekdoty o klíčení doma.
00:43:14 – O teplotách varenia a aktivite sulforafanu.
00:43:45 – Premena sulforafanu z glukorafanínu črevnými baktériami.
00:44:24 – Doplnky fungujú lepšie v kombinácii s aktívnou myrozinázou zo zeleniny.
00:44:56 – Techniky varenia a krížová zelenina.
00:46:06 – Izotiokyanáty ako struma.
Poďakovanie
Túto prácu podporila Fínska akadémia, Nadácia Sigrid Juselius a Fínske organizácie proti rakovine.
Záverom možno povedať, že nukleárny faktor (odvodený od erytroidov 2) podobný 2, tiež známy ako NFE2L2 alebo Nrf2, je proteín, ktorý zvyšuje produkciu antioxidantov, ktoré chránia ľudské telo pred oxidačným stresom. Ako je opísané vyššie, stimulácia dráhy Nrf2 sú štúdie na liečbu chorôb spôsobených oxidačným stresom, vrátane rakoviny. Rozsah našich informácií je obmedzený na chiropraktické a zdravotné problémy chrbtice. Ak chcete prediskutovať túto tému, neváhajte sa opýtať Dr. Jimeneza alebo nás kontaktujte na adrese�915-850-0900 .
Ďalšia téma diskusie: Zmiernenie bolesti kolena bez operácie
Bolesť kolena je dobre známym príznakom, ktorý sa môže vyskytnúť v dôsledku rôznych zranení a/alebo stavov kolena, vrátane�športové zranenia. Koleno je jedným z najzložitejších kĺbov v ľudskom tele, pretože je tvorené priesečníkom štyroch kostí, štyroch väzov, rôznych šliach, dvoch meniskov a chrupavky. Podľa Americkej akadémie rodinných lekárov medzi najčastejšie príčiny bolesti kolena patrí patelárna subluxácia, patelárna tendinitída alebo skokanovo koleno a Osgood-Schlatterova choroba. Hoci bolesť kolena sa s najväčšou pravdepodobnosťou vyskytuje u ľudí starších ako 60 rokov, bolesť kolena sa môže vyskytnúť aj u detí a dospievajúcich. Bolesť kolena sa môže liečiť doma podľa metód RICE, avšak vážne poranenia kolena môžu vyžadovať okamžitú lekársku pomoc vrátane chiropraktickej starostlivosti.
DNA podporuje približne 20,000 XNUMX génov, z ktorých každý obsahuje program na vytvorenie proteínu alebo enzýmu potrebného pre zdravý životný štýl. Každý z týchto vzorcov musí byť neustále regulovaný akýmsi „promótorom“, ktorý presne riadi, koľko z každej látky a/alebo chemikálie sa vytvorí a za akých podmienok sa tiež vyvinie.
Pripojením k určitému druhu oblastí promótora podobných prepínačom, známym ako prvok antioxidačnej odozvy alebo ARE, Faktor Nrf2�podporuje rýchlosť tvorby stoviek odlišných génov, ktoré umožňujú bunkám prežiť v stresujúcich podmienkach. Tieto gény potom generujú výber antioxidačných enzýmov, ktoré vytvárajú obrannú sieť neutralizáciou oxidantov a čistením toxických vedľajších produktov, ktoré zostali pri ich výrobe, navyše pomáhajú obnoviť škody, ktoré spôsobili.
Čo je to oxidačný stres?
Niekoľko oxidantov, ako je superoxidový radikál alebo O2-, a peroxid vodíka alebo H2O2, bolo vytvorených prostredníctvom praxe spaľovania látok a/alebo chemikálií, ktoré udržujú ľudské telo. Ľudské telo má antioxidačné enzýmy, ktoré neutralizujú a detoxikujú reaktívne potraviny a nápoje, ktoré konzumujeme. Nrf2 moduluje ich produkciu, aby sa udržala rovnováha a podčiarkuje dopyt po všetkých týchto enzýmoch. Táto rovnováha môže byť narušená niekoľkými faktormi, vrátane veku.
Ako starneme, ľudské telo vytvára menej Nrf2 a táto jemná rovnováha sa môže postupne začať otáčať smerom k oxidačnej strane, čo je stav označovaný ako oxidačný stres. Choroba môže tiež spôsobiť nadprodukciu oxidantov. Infekcie, alergie a autoimunitné poruchy môžu navyše spustiť naše imunitné bunky, aby vytvorili reaktívne oxidanty, ako je O2-. , H2O2, OH a HOCl, kde sa zdravé bunky poškodia a reagujú zápalom. Choroby spojené so starnutím, vrátane srdcových infarktov, mŕtvice, rakoviny a neurodegeneratívnych stavov, ako je Alzheimerova choroba, tiež zvyšujú vývoj oxidantov, vytvárajú stres a zápalovú reakciu.
Čo sú aktivátory Nrf2?
Proteín Nrf2, tiež nazývaný transkripčný faktor vďaka spôsobu, akým môže podporovať a kontrolovať enzýmy a gény, je tajným prvkom sekvencie biochemických reakcií v bunke, ktorá reaguje na zmeny kognitívnej rovnováhy, ako aj oxidačnej rovnováhy. Snímacie prvky tejto dráhy modifikujú a vybíjajú Nrf2, čím ho spúšťajú, aby sa mohol šíriť do jadra bunky smerom k DNA. Nrf2 môže alternatívne zapnúť alebo vypnúť gény a enzýmy, ktoré podporuje na ochranu bunky.
Našťastie sa rôzne látky, ktoré sú aktivátormi Nrf2, vyvíjajú konzumáciou určitých rastlín a extraktov používaných pred storočiami v čínskych a indiánskych tradičných liekoch. Zdá sa, že tieto fytochemikálie sú rovnako účinné s menším počtom vedľajších účinkov ako farmaceutické produkty aktivujúce Nrf2, ktoré sa dnes používajú.
Faktor súvisiaci s erytroidným faktorom 2, bežnejšie známy ako Nrf2, je transkripčný faktor, ktorý chráni bunku reguláciou génov, enzýmov a antioxidačných reakcií. Transkripčné faktory sú typom proteínu, ktorý sa pripája k DNA, aby podporil tvorbu špecifických látok a chemikálií, vrátane glutatión S-transferáz alebo GST. Aktivácia Nrf2 indukuje produkciu aktívnych proteínov, ktoré vykazujú silnú antioxidačnú kapacitu, ktorá pomáha znižovať oxidačný stres.
Dr. Alex Jimenez DC, CCST Insight
Veda za aktiváciou Nrf2
Po vytvorení prvého výživového doplnku aktivujúceho Nrf2 v roku 2004 bolo známych len minimum informácií o funkcii dráhy Nrf2. V literatúre existovalo približne 200 novín o Nrf2, tiež známych ako nukleárny faktor 2 alebo NFE2L2, a výskumníci ešte len začínali objavovať antioxidačnú reakciu Nrf2 u cicavcov. Od roku 2017 však bolo vytlačených viac ako 9,300 XNUMX akademických výskumných štúdií o tomto „hlavnom regulátore“.
V skutočnosti Nrf2 reguluje mnohé antioxidačné enzýmy, ktoré nesúvisia s génmi, namiesto toho ponúkajú ochranu pred rôznymi okolnosťami súvisiacimi so stresom, s ktorými sa stretávajú bunky, orgány a nakoniec organizmy za zdravých a patologických podmienok. Na základe tohto nového množstva informácií z publikovaných akademických výskumných štúdií sa teraz výskumníci môžu lepšie rozvíjať Výživové doplnky Nrf2.
Od roku 2007 výskumné štúdie preukázali komplexnú funkciu dráhy Nrf2. Zistilo sa, že aktivátory Nrf2 napodobňujú faktory rôznych štruktúr v ľudskom tele. Prostredníctvom týchto dráh boli aktivátory Nrf2 vybavené tak, aby cítili meniace sa podmienky v celej bunke, aby udržali rovnováhu a reagovali na vyvíjajúce sa požiadavky génov.
Prečo používať doplnky aktivujúce Nrf2?
Keď sa schopnosti aktivácie Nrf2 s vekom v organizmoch zmenšujú, môžu sa začať objavovať zmeny. Výskumné štúdie ukázali, že zameranie Nrf2 v bunkách s vekom klesá, čo ukazuje na zvýšené markery oxidačného stresu. V dôsledku týchto zmien sa môžu vyvinúť rôzne choroby súvisiace s vekom, ako je ateroskleróza a kardiovaskulárne choroby, artritída, rakovina, obezita, diabetes typu 2, hypertenzia, šedý zákal a Alzheimerova choroba, ako aj Parkinsonova choroba. Pri týchto zdravotných problémoch sa zistil oxidačný stres.
Stimuláciou bunkovej kapacity zvýšiť produkciu aktivátorov Nrf2, Výživové doplnky Nrf2 môže pomôcť oživiť vlastnú schopnosť ľudského tela pôsobiť proti účinkom oxidačného stresu. Polynenasýtené mastné kyseliny alebo PUFA sú jednou z najľahšie oxidovaných molekúl a sú obzvlášť náchylné na poškodenie voľnými radikálmi. Produkcia kyseliny tiobarbiturovej alebo TBARS sa môže s vekom zvyšovať, čo naznačuje zvýšený oxidačný stres spolu s poklesom dráh regulovaných Nrf2.
Z biologického hľadiska je génová indukcia skutočne pomalým mechanizmom, ktorý si vo všeobecnosti vyžaduje hodiny na prenos cestou. Výsledkom je, že mnohé enzýmy majú svoje vlastné vypínače, ktoré môžu byť spustené v priebehu niekoľkých minút rôznymi regulačnými enzýmami. Výskumníci vyvinuli patentované kompozície aktivátorov Nrf2, ktoré využívajú túto znalostnú bázu aktivácie. Aktivácia Nrf2 pozostáva nielen z transkripčného faktora Nrf2, ktorý sa uvoľňuje z jeho inhibítora a migruje do bunkového jadra, ale tiež sa viaže na špecifické sekvencie DNA, aby sa podporila expresia cytoprotektívneho génu, čím sa reguluje rýchlosť, pri ktorej sa Nrf2 vyberie z jadra.
Pochopenie postupu eliminácie a aktivácie Nrf2 v ľudskom tele umožnilo výskumníkom vytvoriť kombinácie rôznych aktivátorov Nrf2 na dosiahnutie odrazu génov prostredníctvom jeho modulácie. Kombinácia vedomostnej základne spolu so širokou škálou ďalších výskumných štúdií pomohla vyrobiť aktivátory Nrf2 na použitie ako doplnky stravy. Rozsah našich informácií je obmedzený na chiropraktické a zdravotné problémy chrbtice. Ak chcete prediskutovať túto tému, neváhajte sa opýtať Dr. Jimeneza alebo nás kontaktujte na adrese�915-850-0900 .
Kurátorom je Dr. Alex Jimenez
Ďalšia téma diskusie: Zmiernenie bolesti kolena bez operácie
Bolesť kolena je dobre známym príznakom, ktorý sa môže vyskytnúť v dôsledku rôznych zranení a/alebo stavov kolena, vrátane�športové zranenia. Koleno je jedným z najzložitejších kĺbov v ľudskom tele, pretože je tvorené priesečníkom štyroch kostí, štyroch väzov, rôznych šliach, dvoch meniskov a chrupavky. Podľa Americkej akadémie rodinných lekárov medzi najčastejšie príčiny bolesti kolena patrí patelárna subluxácia, patelárna tendinitída alebo skokanovo koleno a Osgood-Schlatterova choroba. Hoci bolesť kolena sa s najväčšou pravdepodobnosťou vyskytuje u ľudí starších ako 60 rokov, bolesť kolena sa môže vyskytnúť aj u detí a dospievajúcich. Bolesť kolena sa môže liečiť doma podľa metód RICE, avšak vážne poranenia kolena môžu vyžadovať okamžitú lekársku pomoc vrátane chiropraktickej starostlivosti.
Antioxidanty sú vedecky označované ako zlúčeniny, ktoré obmedzujú oxidačný proces v ľudskom tele, ktorý ak nie je kontrolovaný, môže vytvárať voľné radikály, ktoré môžu vyvolať početné reťazové reakcie, ktoré môžu spôsobiť poškodenie buniek. Našťastie si ľudské telo dokáže vytvoriť takéto vstavané imunitné mechanizmy, ale keď sa hromadia reaktívne formy kyslíka alebo ROS, nie sú schopné neutralizovať, predstavte si malý plameň, ktorý sa vymkne kontrole, keď sa naplní kyslíkom, nevyhnutne dôjde k poškodeniu. .
Aby sme pokračovali v rozširovaní metafory plameňa, konečným produktom neschopnosti neutralizovať vplyv ROS alebo reaktívnych foriem kyslíka je poškodenie a zápal, inými slovami, ľudské telo doslova horí. Fantastická vec je, že existujú antioxidanty, ktoré môžu ohromne pomôcť v boji proti tomuto zdravotnému problému, a týmto antioxidantom je glutatión. Hoci sa antioxidačný účinok glutatiónu objavil v roku 1889, stal sa jednou z najzaujímavejších tém v moderných výskumných štúdiách.
Majster antioxidantov: glutatión
Silnou látkou je tripeptid, ktorý sa vyvíja z cysteínu, kyseliny glutámovej a glycínu. Vďaka svojej schopnosti chrániť ľudské telo pred tvorbou voľných radikálov môže glutatión v konečnom dôsledku prispieť k podpore zdravého imunitného systému. Založené na Vedecké správy za rok 2015sa zistilo, že schopnosť glutatiónu fungovať synergicky s peroxiredínom a katalázou pomáha chrániť bunky pred peroxidom vodíka. Tento synergický vzorec pôsobí proti reaktívnym druhom kyslíka alebo ROS. Glutatión, peroxidredín a kataláza sú základnými prvkami pri zvyšovaní bunkovej homeostázy, čo je základný proces zdravých buniek, tkanív a orgánov ako celku.
Okrem toho glutatión zvyšuje celkovú štruktúru a funkciu imunitného systému s využitím jeho dôležitého účinku na funkcie lymfocytov. Podľa Ústav imunochémieSprávne dopĺňanie hladín glutatiónu v ľudskom tele môže výrazne posilniť imunitné reakcie. Napríklad dve randomizované placebom kontrolované štúdie ukázali, že terapeutická liečba pacientov s oslabeným imunitným systémom N-acetylcysteínom alebo NAC viedla v oboch prípadoch k podstatnému nárastu väčšiny imunologických procesov, ktoré zahŕňali úplné omladenie. aktivity prirodzených zabíjačských buniek. N-acetyl-cysteín alebo NAC využíva síru z glutatiónu a spája ju s jedovatými molekulami, ktoré sa potom stávajú rozpustnými vo vode a vylučujú sa v ľudskom tele.
Glutatión má tiež schopnosť revitalizovať kyselinu lipoovú, ako aj recyklovať vitamín C a E, ktoré sú potrebné na spustenie určitých systémových procesov vyslaním elektrónov na neutralizáciu voľných radikálov. Na základe výskumnej štúdie z PLoS ONEglutatiónom postihnutí pacienti s diabetes metillus alebo T2DM a mycobacterium tuberculosis. Normálne majú jedinci so slabým imunitným systémom tendenciu vykazovať väčšiu expozíciu M. tb alebo mycobacterium tuberculosis, chorobe alebo infekcii. Okrem toho, jedinci s diabetes metilllus typu 2 alebo T2DM sú dvakrát až trikrát náchylnejší na TBC ako ľudia bez T2DM. Výskumná štúdia tiež naznačila, že zvýšenie hladín glutatiónu v makrofágoch izolovaných od pacientov s T2DM viedlo k zlepšeniu kontroly ochorenia alebo infekcie M.Tb. Tieto výsledky ukazujú, že nižšie hladiny glutatiónu u pacientov s T2DM prispievajú k zvýšenému riziku ochorenia alebo infekcie M. tb. Navyše v závislosti od Dietro Ghezzi v Brightone a Sussex Medical SchoolOxidačný stres môže v konečnom dôsledku spôsobiť zlú štruktúru a funkciu imunitného systému.
Našťastie glutatión hrá zásadnú úlohu pri posilňovaní a kontrole imunity. Napríklad glutatión je nevyhnutný pre vrodené a adaptačné procesy v rámci imunitného systému, vrátane proliferácie T-lymfocytov, fagocytárnej aktivity polymorfonukleárnych neutrofilov a funkcií dendritických buniek, ktoré môžu byť zásadné, pretože sa skladajú z buniek prezentujúcich antigén. . Bunkami sprostredkovaná imunita zahŕňa proteínové antigény, ktoré spočiatku začínajú degenerovať v endocytových vezikulách makrofágov a dendritických buniek, a preto je na povrchu preukázané, že menšie peptidy aktivujú proliferáciu antigén-špecifických T buniek. Okrem toho glutatión pomáha pri tvorbe cytokínov a je potrebné udržiavať produkciu interferónu-gama dendritickými bunkami, čo je dôležité pri ochrane pred vnútrobunkovými patogénmi vrátane mykobaktérií.
N-acetyl-cysteín alebo NAC, vedecky označovaný ako prekurzor glutatiónu, je tiež veľmi silný bunkový antioxidant používaný ako antioxidant zachytávajúci voľné radikály. Všeobecne uznávaná pre svoju úlohu v odvrátenie toxicity acetaminofénu, NAC alebo N-acetyl-cysteín, preukázalo sa, že má niekoľko zdravotných a wellness výhod. Podľa Cell JournalNAC pomáha podporovať zdravú zápalovú reakciu a môže mať pozitívny vplyv na ľudský termín a predčasný pôrod. Výskumná štúdia dospela k záveru, že u žien s predchádzajúcim predčasným pôrodom a bakteriálnou vaginózou, 0.6 gramu NAC denne užívaného perorálne spolu s progesterónom po 16. týždni tehotenstva chránilo pred recidívou predčasného pôrodu a zlepšilo novorodenecký výsledok. Záverom možno povedať, že boli zistené aj pozitívne účinky NAC na budovanie svalov. Po troch minútach pretrvávajúcich kontrakcií došlo k 15-percentnému zvýšeniu výdaja, čo dokazuje, ako NAC hrá zásadnú úlohu pri zlepšovaní budovania svalov a znižovaní celkovej únavy počas pôrodu.
Výskumníci tiež zistili, že NAC alebo N-acetyl-cysteín môže byť prínosom pre tých, ktorí majú syndróm polycystických vaječníkov alebo PCOS. PCOS alebo syndróm polycystických ovárií je bežné ochorenie súvisiace s endokrinnými žľazami, ktoré postihuje približne 5 až 10 percent žien v reprodukčnom veku. U takýchto pacientov existuje väčšie riziko výskytu metabolického syndrómu, kde použitie NAC pomohlo obnoviť zdravé hladiny inzulínu a citlivosť.
Pohľad Dr. Alexa Jimeneza
Glutatión je označovaný ako „majster antioxidantov“ kvôli jeho základnej úlohe pri dosahovaní a udržiavaní celkového zdravia a pohody. Zatiaľ čo ľudské telo je schopné produkovať svoj vlastný glutatión, zlá výživa, znečistenie, toxíny, nadmerné užívanie drog a/alebo liekov, stres, trauma, starnutie, choroby a radiácia, to všetko môže znížiť naše prirodzené hladiny glutatiónu. To môže následne spôsobiť, že jednotlivci sú náchylnejší na poškodenie buniek oxidačným stresom, voľnými radikálmi, infekciami a rakovinou. Suplementácia glutatiónu preto môže mať pre ľudský organizmus obrovské výhody. Spolu s alternatívnymi možnosťami liečby, ako je chiropraktická starostlivosť, možno hladiny glutatiónu opäť regulovať, aby sa zlepšila pohoda.
Zdravotnícki pracovníci navyše navrhli zaviesť užívanie suplementácie glutatiónu spolu s ďalšími alternatívnymi možnosťami liečby, ako napr chiropraxe starostlivostina ďalšie zlepšenie celkového zdravia a pohody. Antioxidanty sú dôležité na udržanie maximálnej pohody, ako aj na inhibíciu reťazovej reakcie voľných radikálov, ktoré spôsobujú poškodenie alebo poškodenie buniek. Výkonné antioxidanty ako glutatión, ako už bolo spomenuté vyššie, v konečnom dôsledku pomáhajú regulovať vývoj týchto voľných radikálov a poskytujú zdravšiu odpoveď imunitného systému. Výskumné štúdie to zistili chiropraxe starostlivosti môže tiež zohrávať podstatnú úlohu v tomto procese, prirodzene zvyšuje aktivitu antioxidantov v ľudskom tele. Chiropraktická starostlivosť je bezpečný a účinný liečebný prístup, ktorý využíva úpravy chrbtice a ručné manipulácie na korekciu vychýlenia chrbtice alebo subluxácií, aby sa ľudskému telu umožnilo prirodzené vyliečenie bez použitia liekov/liekov a/alebo chirurgických zákrokov.
Napokon, antioxidanty demonštrujú svoje biologické vlastnosti prostredníctvom veľkého množstva zdravotných výhod, ktoré môžu byť teraz potrebné viac ako kedykoľvek predtým s každým tak rastúcim náporom stresu, chorôb a znečistenia v našom modernom svete, ktoré všetky prispievajú k poškodeniu a/alebo poškodeniu buniek. . Glutatión a jeho prekurzor NAC alebo N-acetyl-cysteín naďalej preukazujú svoje silné postavenie v oblasti antioxidantov. Spolu s alternatívnymi možnosťami liečby, ako je chiropraktická starostlivosť, môžu ľudia využiť všetky výhody, ktoré tento silný antioxidant ponúka. Rozsah našich informácií je obmedzený na chiropraktiku, ako aj na poranenia a stavy chrbtice. Ak chcete prediskutovať túto tému, neváhajte sa opýtať Dr. Jimeneza alebo nás kontaktujte na adrese�915-850-0900 .
Kurátorom je Dr. Alex Jimenez
Ďalšie témy: Bolesť chrbta
Bolesti chrbta je jednou z najčastejších príčin invalidity a vymeškaných dní v práci na celom svete. Bolesti chrbta sa v skutočnosti pripisujú ako druhý najčastejší dôvod návštevy lekára, prevyšujú ju len infekcie horných dýchacích ciest. Približne 80 percent populácie aspoň raz za život zažije nejaký typ bolesti chrbta. Chrbtica je zložitá štruktúra pozostávajúca z kostí, kĺbov, väzov a svalov, okrem iného z mäkkých tkanív. Z tohto dôvodu môžu zranenia a/alebo zhoršené stavy, ako napr herné disky, môže nakoniec viesť k príznakom bolesti chrbta. Športové zranenia alebo zranenia pri automobilových nehodách sú často najčastejšou príčinou bolesti chrbta, niekedy však môžu mať aj tie najjednoduchšie pohyby bolestivé následky. Našťastie alternatívne možnosti liečby, ako je chiropraktická starostlivosť, môžu pomôcť zmierniť bolesť chrbta pomocou úprav chrbtice a manuálnych manipulácií, čo v konečnom dôsledku zlepšuje úľavu od bolesti.
Vedecky založený chiropraktik Dr. Alexander Jimenez sa pozrie na oxidačný stres, čo to je, ako to ovplyvňuje telo a antioxidačnú obranu na nápravu situácie.
Esra Birben PhD, 1 Umit Murat Sahiner MD, 1 Cansin Sackesen MD, 1 Serpil Erzurum MD, 2 a Omer Kalayci, MD1
Abstrakt: Reaktívne formy kyslíka (ROS) sú produkované živými organizmami v dôsledku normálneho bunkového metabolizmu a environmentálnych faktorov, ako sú látky znečisťujúce ovzdušie alebo cigaretový dym. ROS sú vysoko reaktívne molekuly a môžu poškodiť bunkové štruktúry, ako sú sacharidy, nukleové kyseliny, lipidy a proteíny, a zmeniť ich funkcie. Posun v rovnováhe medzi oxidantmi a antioxidantmi v prospech oxidantov sa nazýva �oxidačný stres.� Regulácia redukčného a oxidačného (redoxného) stavu je rozhodujúca pre životaschopnosť buniek, aktiváciu, proliferáciu a funkciu orgánov. Aeróbne organizmy majú integrované antioxidačné systémy, ktoré zahŕňajú enzymatické a neenzymatické antioxidanty, ktoré sú zvyčajne účinné pri blokovaní škodlivých účinkov ROS. Pri patologických stavoch však môžu byť antioxidačné systémy preťažené. Oxidačný stres prispieva k mnohým patologickým stavom a ochoreniam, vrátane rakoviny, neurologických porúch, aterosklerózy, hypertenzie, ischémie/perfúzie, cukrovky, syndrómu akútnej respiračnej tiesne, idiopatickej pľúcnej fibrózy, chronickej obštrukčnej choroby pľúc a astmy. V tomto prehľade sumarizujeme bunkové oxidačné a antioxidačné systémy a diskutujeme o bunkových účinkoch a mechanizmoch oxidačného stresu.
Kľúčové slová: antioxidant, oxidant, oxidačný stres, reaktívne formy kyslíka, redox
(WAO Journal 2012; 5:9–19)
Reaktívne formy kyslíka (ROS) sú produkované živými organizmami ako výsledok normálneho bunkového metabolizmu. Pri nízkych až stredných koncentráciách fungujú vo fyziologických bunkových procesoch, ale pri vysokých koncentráciách spôsobujú nepriaznivé modifikácie bunkových komponentov, ako sú lipidy, proteíny a DNA.1�6 Posun v rovnováhe medzi oxidantom/antioxidantom v prospech oxidantov sa nazýva �oxidačný stres.� Oxidačný stres prispieva k mnohým patologickým stavom, vrátane rakoviny, neurologických porúch,7�10 aterosklerózy, hypertenzie, ischémie/perfúzie,11�14 cukrovky, syndrómu akútnej respiračnej tiesne, idiopatickej pľúcnej fibrózy, chronickej obštrukčnej choroby pľúc ,15 a astma.16�21 Aeróbne organizmy majú integrované antioxidačné systémy�, ktoré zahŕňajú enzymatické a neenzymatické antioxidanty, ktoré sú zvyčajne účinné pri blokovaní škodlivých účinkov ROS. Pri patologických stavoch však môžu byť antioxidačné systémy preťažené. V tomto prehľade sumarizujeme bunkové oxidačné a antioxidačné systémy a reguláciu redukčného a oxidačného (redoxného) stavu pri zdravotných a chorobných stavoch.
OXIDANTY
Endogénne zdroje ROS
ROS sú produkované z molekulárneho kyslíka ako výsledok normálneho bunkového metabolizmu. ROS možno rozdeliť do 2 skupín: voľné radikály a neradikály. Molekuly obsahujúce jeden alebo viac nepárových elektrónov, ktoré molekule poskytujú reaktivitu, sa nazývajú voľné radikály. Keď 2 voľné radikály zdieľajú svoje nepárové elektróny, vytvoria sa neradikálové formy. Tri hlavné ROS, ktoré majú fyziologický význam, sú superoxidový anión (O3.), hydroxylový radikál (OH) a peroxid vodíka (H22O2). ROS sú zhrnuté v tabuľke 2.
Superoxidový anión vzniká adíciou 1 elektrónu k molekulárnemu kyslíku.22 Tento proces sprostredkúva nikotín adenín dinukleotid fosfát [NAD(P)H] oxidáza alebo xantín oxidáza alebo mitochondriálny elektrónový transportný systém. Hlavným miestom na produkciu superoxidového aniónu sú mitochondrie, mechanizmus bunky na produkciu adenozíntrifosfátu. Normálne sa elektróny prenášajú cez mitochondriálny elektrónový transportný reťazec na redukciu kyslíka na vodu, ale približne 1 až 3 % všetkých elektrónov uniká zo systému a produkuje superoxid. NAD(P)H oxidáza sa nachádza v polymorfonukleárnych leukocytoch, monocytoch a makrofágoch. Po fagocytóze tieto bunky produkujú výbuch superoxidu, ktorý vedie k baktericídnej aktivite. Superoxid sa premieňa na peroxid vodíka pôsobením superoxiddismutáz (SODs, EC 1.15.1.1). Peroxid vodíka ľahko difunduje cez plazmatickú membránu. Peroxid vodíka je tiež produkovaný xantínoxidázou, oxidázou aminokyselín a NAD(P)H oxidázou�23,24 a v peroxizómoch spotrebou molekulárneho kyslíka v metabolických reakciách. V sérii reakcií nazývaných Haber�Weissove a Fentonove reakcie sa H2O2 môže rozložiť na OH2 v prítomnosti transmisných kovov ako Fe21 alebo Cu21.25
Fe31 +�.O2�?Fe2 +�O2 Haber Weiss
Fe2 +�H2O2�?Fe3 +�OH�+ .OH Fentonova reakcia
Samotný O 2 môže tiež reagovať s H2 O2 a vytvárať OH. 26,27 Hydroxylový radikál je najreaktívnejší z ROS a môže poškodiť proteíny, lipidy, sacharidy a DNA. Môže tiež spustiť peroxidáciu lipidov tým, že vezme elektrón z polynenasýtených mastných kyselín.
Granulocytové enzýmy ďalej rozširujú reaktivitu H2O2 prostredníctvom eozinofilnej peroxidázy a myeloperoxidázy (MPO). V aktivovaných neutrofiloch je H2O2 spotrebovaný MPO. V prítomnosti chloridového iónu sa H2O2 premení na kyselinu chlórnu (HOCl). HOCl je vysoko oxidačný a hrá dôležitú úlohu pri ničení patogénov v dýchacích cestách.28 HOCl však môže tiež reagovať s DNA a indukovať interakcie DNA�proteínu a produkovať pyrimidínové oxidačné produkty a pridávať chloridy k bázam DNA.29,30 Eozinofilná peroxidáza a MPO tiež prispievajú k oxidačnému stresu modifikáciou proteínov halogenáciou, nitráciou a zosieťovaním proteínov prostredníctvom tyrozylových radikálov.31�33
Ďalšie voľné radikály odvodené od kyslíka sú peroxylové radikály (ROO$). Najjednoduchšou formou týchto radikálov je hydroperoxylový radikál (HOO$) a zohráva úlohu pri peroxidácii mastných kyselín. Voľné radikály môžu spustiť reťazové reakcie peroxidácie lipidov abstrahovaním atómu vodíka z bočného reťazca metylénového uhlíka. Lipidový radikál potom reaguje s kyslíkom za vzniku peroxylového radikálu. Peroxylový radikál spúšťa reťazovú reakciu a premieňa polynenasýtené mastné kyseliny na lipidové hydroperoxidy. Lipidové hydroperoxidy sú veľmi nestabilné a ľahko sa rozkladajú na sekundárne produkty, ako sú aldehydy (ako je 4-hydroxy-2,3-nonenal) a malondialdehydy (MDA). Izoprostány sú ďalšou skupinou produktov peroxidácie lipidov, ktoré vznikajú peroxidáciou kyseliny arachidónovej a tiež sa zistilo, že sú zvýšené v plazmových a dychových kondenzátoch astmatikov.34,35 Peroxidácia lipidov narúša integritu bunkových membrán a vedie k preskupeniu membránová štruktúra.
Peroxid vodíka, superoxidový radikál, oxidovaný glutatión (GSSG), MDA, izoprostány, karbonyly a nitrotyrozín možno ľahko merať zo vzoriek plazmy, krvi alebo bronchoalveolárnej laváže ako biomarkery oxidácie štandardizovanými testami.
Exogénny zdroj oxidantov
Cigaretový dym
Cigaretový dym obsahuje veľa oxidantov a voľných radikálov a organických zlúčenín, ako sú superoxid a oxid dusnatý.36 Okrem toho vdychovanie cigaretového dymu do pľúc tiež aktivuje niektoré endogénne mechanizmy, ako je akumulácia neutrofilov a makrofágov, ktoré ešte viac zvyšujú oxidačné poškodenie. .
Vystavenie ozónu
Expozícia ozónu môže spôsobiť peroxidáciu lipidov a vyvolať prílev neutrofilov do epitelu dýchacích ciest. Krátkodobá expozícia ozónu tiež spôsobuje uvoľnenie zápalových mediátorov, ako sú MPO, eozinofilné katiónové proteíny a tiež laktátdehydrogenáza a albumín.37 Dokonca aj u zdravých jedincov spôsobuje expozícia ozónu zníženie pľúcnych funkcií.38 Cho et al39 ukázali, že častice (zmes pevných častíc a kvapiek kvapaliny suspendovaných vo vzduchu) katalyzuje redukciu kyslíka.
Hyperoxia
Hyperoxia sa vzťahuje na stavy s vyššími hladinami kyslíka ako je normálny parciálny tlak kyslíka v pľúcach alebo iných telesných tkanivách. Vedie k väčšej produkcii reaktívnych foriem kyslíka a dusíka.40,41
Ionizujúce žiarenie
Ionizujúce žiarenie v prítomnosti O2 premieňa hydroxylové radikály, superoxidy a organické radikály na peroxid vodíka a organické hydroperoxidy. Tieto hydroperoxidové druhy reagujú s redoxne aktívnymi kovovými iónmi, ako je Fe a Cu, prostredníctvom Fentonových reakcií, a tak indukujú oxidačný stres.42,43 Narayanan et al44 ukázali, že fibroblasty, ktoré boli vystavené alfa časticiam, mali významné zvýšenie intracelulárneho O2. a H2O2 produkcia prostredníctvom NADPH oxidázy viazanej na plazmatickú membránu.2 Molekuly prenosu signálu, ako je extracelulárna signálom regulovaná kináza 44 a 1 (ERK2/1), c-Jun N-terminálna kináza (JNK) a p2, a transkripčné faktory, ako napr. aktivátorový proteín-38 (AP-1), jadrový faktor-kB (NF-kB) a p1, sú aktivované, čo vedie k expresii génov súvisiacich s radiačnou odpoveďou.53�45 Ultrafialové A (UVA) fotóny spúšťajú oxidačné reakcie excitáciou endogénnych fotosenzibilizátorov, ako sú porfyríny, NADPH oxidáza a riboflavíny. 50-Oxo-8-dihydroguanín (7,8-oxoGua) je hlavný UVA-sprostredkovaný produkt oxidácie DNA, ktorý vzniká oxidáciou OH radikálu, 8-elektrónových oxidantov a singletového kyslíka, ktorý reaguje hlavne s guanínom.1 Vznik guanínu Ukázalo sa, že radikálový katión v izolovanej DNA sa účinne vyskytuje prostredníctvom priameho účinku ionizujúceho žiarenia.51 Po vystavení ionizujúcemu žiareniu vnútrobunková hladina glutatiónu (GSH) krátkodobo klesá, ale potom sa opäť zvyšuje.52,53
Ióny ťažkých kovov
Ióny ťažkých kovov, ako je železo, meď, kadmium, ortuť, nikel, olovo a arzén, môžu vyvolať tvorbu reaktívnych radikálov a spôsobiť poškodenie buniek prostredníctvom deplécie enzýmových aktivít prostredníctvom peroxidácie lipidov a reakcie s jadrovými proteínmi a DNA.55
Jedným z najdôležitejších mechanizmov vytvárania voľných radikálov sprostredkovaných kovom je reakcia Fentonovho typu. Superoxidový ión a peroxid vodíka môžu interagovať s prechodnými kovmi, ako je železo a meď, prostredníctvom kovom katalyzovanej Haber�Weiss/Fentonovej reakcie za vzniku OH radikálov.
Okrem mechanizmov Fentonovho typu a Haber-Weissovho typu môžu určité kovové ióny reagovať priamo s bunkovými molekulami za vzniku voľných radikálov, ako sú tiolové radikály, alebo indukovať bunkové signálne dráhy. Tieto radikály môžu tiež reagovať s inými molekulami tiolu za vzniku O22..O22. sa premieňa na H2O2, čo spôsobuje ďalšiu tvorbu kyslíkových radikálov. Niektoré kovy, ako napríklad arzenit, indukujú tvorbu ROS nepriamo aktiváciou systémov produkujúcich radikály v bunkách.56
Arzén je vysoko toxický prvok, ktorý produkuje rôzne ROS, vrátane superoxidu (O2 2), singletového kyslíka (1O2), peroxylového radikálu (ROO ), oxidu dusnatého (NO ), peroxidu vodíka (H2O2) a dimetylarzínových peroxylových radikálov [( CH3)2AsOO ].57�59 Zlúčeniny arzénu (III) môžu inhibovať antioxidačné enzýmy, najmä enzýmy závislé od GSH, ako sú glutatión-S-transferázy (GST), glutatiónperoxidáza (GSH-Px) a GSH reduktáza prostredníctvom väzby - ing na ich sulfhydrylové (�SH) skupiny.60,61
Olovo zvyšuje peroxidáciu lipidov.62 Po expozícii olovom boli zaznamenané významné zníženia aktivity tkanivovej SOD a GPx mozgu.63,64 Nahradenie zinku, ktorý slúži ako kofaktor mnohých enzýmov, olovom, vedie k inaktivácii takýchto enzýmov. Expozícia olova môže spôsobiť inhibíciu GST ovplyvnením tkanivových tiolov.
ROS generované reakciami katalyzovanými kovom môže modifikovať bázy DNA. Tri substitúcie báz, G/C, G/T a C/T, sa môžu vyskytnúť v dôsledku oxidačného poškodenia kovovými iónmi, ako sú Fe21, Cu21 a Ni21. Reid et al65 ukázali, že G/C bol produkovaný prevažne Fe21, zatiaľ čo C/T substitúcia bola Cu21 a Ni21.
ANTIOXIDANTY
Ľudské telo je vybavené množstvom antioxidantov, ktoré slúžia na vyváženie účinku oxidantov. Pre všetky praktické účely ich možno rozdeliť do 2 kategórií: enzymatické (tabuľka 2) a neenzymatické (tabuľka 3).
Enzymatické antioxidanty
Hlavné enzymatické antioxidanty pľúc sú SOD (EC 1.15.1.11), kataláza (EC 1.11.1.6) a GSH-Px (EC 1.11.1.9). Okrem týchto hlavných enzýmov, ďalšie antioxidanty, vrátane hemoxygenázy-1 (EC 1.14.99.3) a redoxných proteínov, ako sú tioredoxíny (TRX, EC 1.8.4.10), peroxiredoxíny (PRX, EC 1.11.1.15) a glutaredoxíny Zistilo sa tiež, že hrajú kľúčovú úlohu v pľúcnej antioxidačnej obrane.
Pretože superoxid je primárny ROS produkovaný z rôznych zdrojov, jeho dismutácia pomocou SOD má primárny význam pre každú bunku. Všetky 3 formy SOD, to znamená CuZn-SOD, Mn-SOD a EC-SOD, sú široko exprimované v ľudských pľúcach. Mn-SOD je lokalizovaný v matrici mitochondrií. EC-SOD je primárne lokalizovaný v extracelulárnej matrici, najmä v oblastiach obsahujúcich veľké množstvo kolagénových vlákien typu I a okolo pľúcnych a systémových ciev. Zistilo sa to aj v bronchiálnom epiteli, alveolárnom epiteli a alveolárnych makrofágoch.66,67 Celkovo sa všeobecne predpokladá, že CuZn-SOD a Mn-SOD pôsobia ako veľké lapače superoxidových radikálov. Relatívne vysoká hladina EC-SOD v pľúcach s jej špecifickou väzbou na zložky extracelulárnej matrice môže predstavovať základnú zložku ochrany pľúcnej matrice.68
H2O2, ktorý vzniká pôsobením SOD alebo pôsobením oxidáz, ako je xantínoxidáza, sa katalázou a GSH-Px redukuje na vodu. Kataláza existuje ako tetramér zložený zo 4 rovnakých monomérov, z ktorých každý obsahuje na aktívnom mieste hemovú skupinu. Degradácia H2O2 sa uskutočňuje premenou medzi 2 konformáciami katalázy-ferikatalázy (železo koordinované s vodou) a zlúčeniny I (železo v komplexe s atómom kyslíka). Kataláza tiež viaže NADPH ako redukčný ekvivalent, aby sa zabránilo oxidatívnej inaktivácii enzýmu (tvorbe zlúčeniny II) H2O2, keď sa redukuje na vodu.69
Enzýmy v redoxnom cykle zodpovedné za redukciu H2O2 a lipidových hydroperoxidov (generovaných ako výsledok membránovej lipidovej peroxidácie) zahŕňajú GSH-Pxs.70 GSH-Pxs sú rodinou tetramérnych enzýmov, ktoré obsahujú jedinečnú aminokyselinu selenocysteín v aktívne miesta a používajú tioly s nízkou molekulovou hmotnosťou, ako je GSH, na redukciu H2O2 a peroxidov lipidov na ich zodpovedajúce alkoholy. Boli opísané štyri GSH-Px kódované rôznymi génmi: GSH-Px-1 (bunkový GSH-Px) je všadeprítomný a redukuje peroxidy H2O2 a mastných kyselín, ale nie esterifikované peroxyllipidy.71 Esterifikované lipidy sú redukované membránovo viazaným GSH -Px-4 (fosfolipid hydroperoxid GSH-Px), ktorý môže využívať niekoľko rôznych nízkomolekulárnych tiolov ako redukčné ekvivalenty. GSH-Px-2 (gastrointestinálny GSH-Px) je lokalizovaný v bunkách gastrointestinálneho epitelu, kde slúži na redukciu peroxidov v potrave.72 GSH-Px-3 (extracelulárny GSH-Px) je jediným členom rodiny GSH-Px, ktorý sídli v extracelulárneho kompartmentu a je považovaný za jeden z najdôležitejších extracelulárnych antioxidačných enzýmov u cicavcov. Z nich sa extracelulárny GSH-Px najviac skúma v ľudských pľúcach.73
Okrem toho je likvidácia H2O2 úzko spojená s niekoľkými enzýmami obsahujúcimi tiol, menovite TRX (TRX1 a TRX2), tioredoxín reduktázy (EC 1.8.1.9) (TRR), PRX (čo sú tioredoxín peroxidázy) a glutaredoxíny.74
V ľudských bunkách boli charakterizované dva TRX a TRR, existujúce v cytosóle aj mitochondriách. V pľúcach sú TRX a TRR exprimované v bronchiálnom a alveolárnom epiteli a makrofágoch. V ľudských bunkách sa našlo šesť rôznych PRX, ktoré sa líšia svojou ultraštrukturálnou kompartmentalizáciou. Experimentálne štúdie odhalili dôležitosť PRX VI pri ochrane alveolárneho epitelu. Ľudské pľúca exprimujú všetky PRX v bronchiálnom epiteli, alveolárnom epiteli a makrofágoch.75 Nedávno sa zistilo, že PRX V funguje ako peroxynitritreduktáza,76 čo znamená, že môže fungovať ako potenciálna ochranná zlúčenina pri rozvoji poškodenia pľúc sprostredkovaného ROS .77
Pre tieto antioxidanty je spoločná požiadavka NADPH ako redukčného ekvivalentu. NADPH udržuje katalázu v aktívnej forme a používa sa ako kofaktor TRX a GSH reduktázy (EC 1.6.4.2), ktorá premieňa GSSG na GSH, ko-substrát pre GSH-Pxs. Intracelulárny NADPH sa zase vytvára redukciou NADP1 glukózo-6-fosfátdehydrogenázou, prvým a rýchlosť obmedzujúcim enzýmom pentózofosfátovej dráhy, počas konverzie glukózo-6-fosfátu na 6-fosfoglukonolaktón. Generovaním NADPH je glukózo-6-fosfátdehydrogenáza kritickým determinantom tlmivej kapacity cytosolického GSH (GSH/GSSG), a preto ju možno považovať za nevyhnutný regulačný antioxidačný enzým.78,79
GST (EC 2.5.1.18), ďalšia skupina antioxidačných enzýmov, inaktivuje sekundárne metabolity, ako sú nenasýtené aldehydy, epoxidy a hydroperoxidy. Boli opísané tri hlavné rodiny GST: cytosolická GST, mitochondriálna GST80,81 a mikrozomálna GST spojená s membránou, ktorá má úlohu v metabolizme eikosanoidov a GSH.82 U cicavcov je identifikovaných sedem tried cytosolických GST, označených ako Alpha, Mu, Pi, Sigma, Theta, Omega a Zeta.83�86 Počas nestresových podmienok GST triedy Mu a Pi interagujú s kinázami Ask1 a JNK a inhibujú tieto kinázy.87�89 Ukázalo sa, že GSTP1 disociuje z JNK ako odpoveď na oxidačný stres.89 GSTP1 tiež fyzicky interaguje s PRX VI a vedie k obnoveniu aktivity enzýmu PRX prostredníctvom glutationylácie oxidovaného proteínu.90
Neenzymatické antioxidanty
Neenzymatické antioxidanty zahŕňajú zlúčeniny s nízkou molekulovou hmotnosťou, ako sú vitamíny (vitamíny C a E), b-karotén, kyselina močová a GSH, tripeptid (Lg-glutamyl-L-cysteinyl-L-glycín), ktorý obsahuje tiol ( sulfhydryl) skupina.
Vitamín C (kyselina askorbová)
Vo vode rozpustný vitamín C (kyselina askorbová) poskytuje intracelulárnu a extracelulárnu antioxidačnú kapacitu vo vodnej fáze predovšetkým zachytávaním voľných kyslíkových radikálov. Premieňa voľné radikály vitamínu E späť na vitamín E. Ukázalo sa, že jeho plazmatické hladiny s vekom klesajú.91,92
Vitamín E (a-tokoferol)
Vitamín E rozpustný v lipidoch je koncentrovaný v hydrofóbnom vnútornom mieste bunkovej membrány a je hlavnou obranou proti poškodeniu membrány vyvolanému oxidantmi. Vitamín E daruje elektrón peroxylovému radikálu, ktorý vzniká pri peroxidácii lipidov. a-Tokoferol je najaktívnejšia forma vitamínu E a hlavný membránovo viazaný antioxidant v bunke. Vitamín E spúšťa apoptózu rakovinových buniek a inhibuje tvorbu voľných radikálov.93
Glutatión
GSH je vysoko hojný vo všetkých bunkových kompartmentoch a je hlavným rozpustným antioxidantom. Pomer GSH/GSSG je hlavným determinantom oxidačného stresu. GSH prejavuje svoje antioxidačné účinky niekoľkými spôsobmi.94 Detoxikuje peroxid vodíka a peroxidy lipidov pôsobením GSH-Px. GSH daruje svoj elektrón H2O2, aby ho redukoval na H2O a O2. GSSG je opäť redukovaný na GSH pomocou GSH reduktázy, ktorá využíva NAD(P)H ako donor elektrónov. GSH-Px sú tiež dôležité pre ochranu bunkovej membrány pred peroxidáciou lipidov. Redukovaný glutatión daruje protóny membránovým lipidom a chráni ich pred útokmi oxidantov.95
GSH je kofaktorom niekoľkých detoxikačných enzýmov, ako sú GSH-Px a transferáza. Má úlohu pri premene vitamínu C a E späť na ich aktívne formy. GSH chráni bunky pred apoptózou interakciou s proapoptotickými a antiapoptotickými signálnymi dráhami.94 Reguluje a aktivuje aj niekoľko transkripčných faktorov, ako sú AP-1, NF-kB a Sp-1.
Karotenoidy (b-karotén)
Karotenoidy sú pigmenty nachádzajúce sa v rastlinách. Predovšetkým sa zistilo, že b-karotén reaguje s peroxylovými (ROO), hydroxylovými (OH) a superoxidovými (O22.) radikálmi.96 Karotenoidy vykazujú svoje antioxidačné účinky pri nízkom parciálnom tlaku kyslíka, ale môžu mať prooxidačné účinky pri vyššom kyslíku. koncentrácií.97 Karotenoidy aj kyseliny retinové (RA) sú schopné regulovať transkripčné faktory.98 b-karotén inhibuje oxidantom indukovanú aktiváciu NF-kB a produkciu interleukínu (IL)-6 a tumor nekrotizujúceho faktora-a. Karotenoidy tiež ovplyvňujú apoptózu buniek. Antiproliferatívne účinky RA boli preukázané v niekoľkých štúdiách. Tento účinok RA je sprostredkovaný hlavne receptormi kyseliny retinovej a líši sa medzi typmi buniek. V bunkách karcinómu prsníka sa ukázalo, že receptor kyseliny retinovej spúšťa inhibíciu rastu indukciou zastavenia bunkového cyklu, apoptózy alebo oboch.99,100
ÚČINOK OXIDATÍVNEHO STRESU: GENETICKÉ, FYZIOLOGICKÉ A BIOCHEMICKÉ MECHANIZMY
Oxidačný stres nastáva, keď je rovnováha medzi antioxidantmi a ROS narušená buď v dôsledku vyčerpania antioxidantov alebo akumulácie ROS. Keď nastane oxidačný stres, bunky sa pokúšajú pôsobiť proti oxidačným účinkom a obnoviť redoxnú rovnováhu aktiváciou alebo umlčaním génov kódujúcich obranné enzýmy, transkripčné faktory a štrukturálne proteíny.101,102 Pomer medzi oxidovaným a redukovaným glutatiónom (2GSH/GSSG) je jeden. z dôležitých determinantov oxidačného stresu v organizme. Vyššia produkcia ROS v tele môže zmeniť štruktúru DNA, viesť k modifikácii proteínov a lipidov, aktivácii viacerých stresom indukovaných transkripčných faktorov a produkcii prozápalových a protizápalových cytokínov.
Účinky oxidačného stresu na DNA
ROS môže viesť k modifikáciám DNA niekoľkými spôsobmi, ktoré zahŕňajú degradáciu báz, jedno- alebo dvojvláknové zlomy DNA, purínové, pyrimidínové alebo na cukre viazané modifikácie, mutácie, delécie alebo translokácie a zosieťovanie s proteínmi. Väčšina týchto modifikácií DNA (obr. 1) je vysoko relevantná pre karcinogenézu, starnutie a neurodegeneratívne, kardiovaskulárne a autoimunitné ochorenia. Tabakový dym, redoxné kovy a neredoxné kovy, ako je železo, kadmium, chróm a arzén, sa tiež podieľajú na karcinogenéze a starnutí tým, že vytvárajú voľné radikály alebo sa viažu na tiolové skupiny. Tvorba 8-OH-G je najznámejším poškodením DNA, ku ktorému dochádza prostredníctvom oxidačného stresu a je potenciálnym biomarkerom karcinogenézy.
Promótorové oblasti génov obsahujú konsenzuálne sekvencie pre transkripčné faktory. Tieto miesta viažuce transkripčný faktor obsahujú sekvencie bohaté na GC, ktoré sú náchylné na oxidačné útoky. Tvorba 8-OH-G DNA vo väzbových miestach pre transkripčný faktor môže modifikovať väzbu transkripčných faktorov a tak zmeniť expresiu príbuzných génov, ako sa ukázalo pre cieľové sekvencie AP-1 a Sp-1 Okrem 103-OH-G, Ukázalo sa tiež, že 8-cyklo-8,59-deoxyadenozín (cyklo-dA) inhibuje transkripciu z reportérového génu v bunkovom systéme, ak sa nachádza v TATA boxe.29 Proteín viažuci TATA iniciuje transkripciu zmenou ohybu DNA. . Väzba proteínu viažuceho TATA môže byť narušená prítomnosťou cyklo-dA.
Oxidačný stres spôsobuje nestabilitu mikrosatelitných (krátke tandemové opakovania) oblastí. Redoxne aktívne kovové ióny, hydroxylové radikály zvyšujú nestabilitu mikrosatelitov.105 Aj keď zlomy jednovláknovej DNA spôsobené oxidačným poškodením môžu bunky ľahko tolerovať, zlomy dvojvláknovej DNA vyvolané ionizujúcim žiarením môžu byť významnou hrozbou pre prežitie bunky.106
Metylácia na CpG ostrovoch v DNA je dôležitý epigenetický mechanizmus, ktorý môže viesť k umlčaniu génov. Oxidácia 5-MeCyt na 5-hydroxymetyluracil (5-OHMeUra) môže prebiehať prostredníctvom deaminačných/oxidačných reakcií tymínu alebo 5-hydroxymetylcytozínových medziproduktov.107 Zdá sa, že okrem modulácie génovej expresie ovplyvňuje organizáciu chromatínu aj metylácia DNA.108 Aberantné vzory metylácie DNA indukované oxidačnými útokmi tiež ovplyvňujú aktivitu opravy DNA.
Účinky oxidačného stresu na lipidy
ROS môže vyvolať peroxidáciu lipidov a narušiť dvojvrstvové usporiadanie membránových lipidov, ktoré môže inaktivovať receptory a enzýmy viazané na membránu a zvýšiť priepustnosť tkanív.109 Produkty peroxidácie lipidov, ako sú MDA a nenasýtené aldehydy, sú schopné inaktivovať mnohé bunkové proteíny vytvorením proteínového kríža. -väzby.110�112 4-Hydroxy-2-nonenal spôsobuje depléciu intracelulárneho GSH a vyvoláva produkciu peroxidu,113,114 aktivuje receptor epidermálneho rastového faktora,115 a indukuje produkciu fibronektínu.116 Produkty peroxidácie lipidov, ako sú izoprostany a látky retiobarbiturovej kyseliny , boli použité ako nepriame biomarkery oxidačného stresu a zvýšené hladiny sa ukázali v kondenzáte vydychovaného dychu alebo tekutine z bronchoalveolárnej laváže alebo pľúcach pacientov s chronickou obštrukčnou chorobou pľúc alebo fajčiarov.117�119
Účinky oxidačného stresu na bielkoviny
ROS môže spôsobiť fragmentáciu peptidového reťazca, zmenu elektrického náboja proteínov, zosieťovanie proteínov a oxidáciu špecifických aminokyselín, a preto vedie k zvýšenej náchylnosti k proteolýze degradáciou špecifickými proteázami.120 Cysteínové a metionínové zvyšky v proteínoch sú obzvlášť náchylné na oxidáciu.121 Oxidácia sulfhydrylových skupín alebo metionínových zvyškov proteínov spôsobuje konformačné zmeny, rozklad proteínov a degradáciu.8,121�123 Enzýmy, ktoré majú kovy na svojich aktívnych miestach alebo blízko nich, sú obzvlášť citlivejšie na oxidáciu katalyzovanú kovom. Ukázalo sa, že oxidačná modifikácia enzýmov inhibuje ich aktivity.124,125
V niektorých prípadoch môže dôjsť k špecifickej oxidácii proteínov. Napríklad metionín môže byť oxidovaný metionínsulfoxid126 a fenylalanín na o-tyrozín127; sulfhydrylové skupiny môžu byť oxidované za vzniku disulfidových väzieb;128 a karbonylové skupiny môžu byť zavedené do bočných reťazcov proteínov. Gama lúče, kovom katalyzovaná oxidácia, HOCl a ozón môžu spôsobiť tvorbu karbonylových skupín.129
Účinky oxidačného stresu na prenos signálu
ROS môže indukovať expresiu niekoľkých génov zapojených do signálnej transdukcie.1,130 Vysoký pomer GSH/GSSG je dôležitý pre ochranu bunky pred oxidačným poškodením. Narušenie tohto pomeru spôsobuje aktiváciu redox citlivých transkripčných faktorov, ako sú NF-kB, AP-1, jadrový faktor aktivovaných T buniek a hypoxiou indukovateľný faktor 1, ktoré sa podieľajú na zápalovej odpovedi. Aktivácia transkripčných faktorov prostredníctvom ROS je dosiahnutá signálnymi transdukčnými kaskádami, ktoré prenášajú informácie zvonku do vnútra bunky. Receptory tyrozínkinázy, väčšina receptorov rastového faktora, ako je receptor epidermálneho rastového faktora, receptor vaskulárneho endotelového rastového faktora a receptor pre rastový faktor odvodený od krvných doštičiek, proteín tyrozínfosfatázy a serín/treonínkinázy sú cieľmi ROS.131�133 Kinázy regulované extracelulárnym signálom, JNK a p38, ktoré sú členmi rodiny mitogénom aktivovaných proteínkináz a podieľajú sa na niekoľkých procesoch v bunke vrátane proliferácie, diferenciácie a apoptózy, môžu byť tiež regulované oxidantmi.
V podmienkach oxidačného stresu cysteínové zvyšky vo väzbovom mieste DNA c-Jun, niektoré podjednotky AP-1 a inhibičná kB kináza podliehajú reverzibilnej S-glutatiolácii. Uvádza sa, že glutaredoxín a TRX hrajú dôležitú úlohu v regulácii redox-senzitívnych signálnych dráh, ako sú NF-kB a AP-1, mitogénom aktivovaná proteínkináza p38 a JNK.134�137
NF-kB sa môže aktivovať v reakcii na podmienky oxidačného stresu, ako sú ROS, voľné radikály a UV žiarenie.138 Fosforylácia IkB uvoľňuje NF-kB a umožňuje mu vstúpiť do jadra, aby sa aktivovala génová transkripcia.139 Množstvo kináz má bolo hlásené, že fosforyluje IkB na serínových zvyškoch. Tieto kinázy sú cieľom oxidačných signálov na aktiváciu NF-kB.140 Redukčné činidlá zvyšujú väzbu NF-kB na DNA, zatiaľ čo oxidačné činidlá inhibujú väzbu DNA na NF-kB. TRX môže vykonávať 2 opačné účinky pri regulácii NF-kB: v cytoplazme blokuje degradáciu IkB a inhibuje aktiváciu NF-kB, ale zvyšuje väzbu NF-kB DNA v jadre.141 Aktivácia NF-kB prostredníctvom degradácie súvisiacej s oxidáciou IkB vedie k aktivácii niekoľkých génov súvisiacich s antioxidačnou obranou. NF-kB reguluje expresiu niekoľkých génov, ktoré sa podieľajú na imunitnej odpovedi, ako sú IL-1b, IL-6, tumor nekrotizujúci faktor-a, IL-8 a niekoľko adhéznych molekúl.142,143 NF-kB tiež reguluje angiogenézu a proliferáciu a diferenciácia buniek.
AP-1 je tiež regulovaný redoxným stavom. V prítomnosti H2O2 môžu niektoré kovové ióny indukovať aktiváciu AP-1. Zvýšenie pomeru GSH/GSSG zvyšuje väzbu AP-1, zatiaľ čo GSSG inhibuje väzbu DNA AP-1.144 Väzba DNA heterodiméru Fos/Jun sa zvyšuje redukciou jedného konzervovaného cysteínu v doméne viažucej DNA každého z proteíny145, zatiaľ čo väzba DNA AP-1 môže byť inhibovaná GSSG v mnohých typoch buniek, čo naznačuje, že tvorba disulfidových väzieb cysteínovými zvyškami inhibuje väzbu DNA AP-1 Prenos signálu prostredníctvom oxidačného stresu je zhrnutý na obrázku 146,147.
záver
Oxidačný stres môže vzniknúť z nadprodukcie ROS metabolickými reakciami, ktoré využívajú kyslík a posúvajú medzi nimi rovnováhu oxidant/antioxidant stavy v prospech oxidantov. ROS sú produkované bunkovými metabolickými aktivitami a environmentálnymi faktormi, ako sú látky znečisťujúce ovzdušie alebo cigaretový dym. ROS sú vysoko reaktívne molekuly kvôli nespárovaným elektrónom vo svojej štruktúre a reagujú s niekoľkými biologickými makromolekulami v bunke, ako sú sacharidy, nukleové kyseliny, lipidy a proteíny, a menia svoje funkcie. ROS tiež ovplyvňuje expresiu niekoľkých génov upreguláciou redox-senzitívnych transkripčných faktorov a remodeláciou chromatínu prostredníctvom zmeny v acetylácii / deacetylácii histónov. Regulácia redoxného stavu je rozhodujúca pre životaschopnosť buniek, aktiváciu, proliferáciu a funkciu orgánov.
REFERENCIE
1. Valko M, Rhodes CJ, Moncol J, Izakovic M, Mazur M. Voľné radikály, kovy a antioxidanty pri rakovine vyvolanej oxidačným stresom. Chem Biol Interact. 2006;160:1�40.
2. Halliwell B, Gutteridge JMC. Voľné radikály v biológii a medicíne. 3. vyd. New York: Oxford University Press;1999.
3. Marnett LJ. Peroxidácia lipidov a poškodenie DNA malondialdehydom. Mutat Res. 1999;424:83�95.
4. Siems WG, Grune T, Esterbauer H. Tvorba 4-hydroxynonenalu počas ischémie a reperfúzie tenkého čreva potkana. �Life Sci. 1995;57:785�789.
5. Stadtman ER. Úloha druhov oxidantov pri starnutí. Curr Med Chem. 2004;11:1105�1112.
6. Wang MY, Dhingra K, Hittelman WN, Liehr JG, deAndrade M, Li DH. Predpokladané malondialdehydové adukty DNA v tkanivách ľudského prsníka indukované peroxidáciou lipidov. Biomarkery epidemiolu rakoviny Predch. 1996;5:705�710.
7. Jenner P. Oxidačný stres pri Parkinsonovej chorobe. Ann Neurol. 2003;53: S26�S36.
8. Lyras L, Cairns NJ, Jenner A, Jenner P, Halliwell B. Hodnotenie oxidačného poškodenia proteínov, lipidov a DNA v mozgu od pacientov s Alzheimerovou chorobou. J Neurochem. 1997;68:2061�2069.
9. Sayre LM, Smith MA, Perry G. Chémia a biochémia oxidačného stresu pri neurodegeneratívnom ochorení. Curr Med Chem. 2001;8:721�738.
10. Toshniwal PK, Zarling EJ. Dôkaz zvýšenej peroxidácie lipidov pri roztrúsenej skleróze. Neurochem Res. 1992;17:205�207.
11. Dhalla NS, Temsah RM, Netticadan T. Úloha oxidačného stresu pri kardiovaskulárnych ochoreniach. J Hypertens. 2000;18:655�673.
12. Kašparová S, Brezová V, Valko M, Horecký J, Mlynárik V, et al. Štúdia oxidačného stresu na potkanom modeli chronickej hypoperfúzie mozgu. Neurochem Int. 2005;46:601�611.
13. Kerr S, Brosnan MJ, McIntyre M, Reid JL, Dominiczak AF, Hamilton CA. Produkcia superoxidových aniónov je zvýšená v modeli genetickej hypertenzie: úloha endotelu. Hypertenzia. 1999;33:1353�1358.
14. Kukreja RC, Hess ML. Systém voľných radikálov kyslíka: od rovníc cez interakcie membránových proteínov až po kardiovaskulárne poškodenie a ochranu. Cardiovasc Res. 1992;26:641�655.
15. Asami S, Manabe H, Miyake J, Tsurudome Y, Hirano T, a kol. Fajčenie cigariet vyvoláva zvýšenie oxidačného poškodenia DNA, 8-hydroxydeoxyguanozín, v centrálnom mieste ľudských pľúc. Karcinogenéza. 1997;18:1763�1766.
16. Andreadis AA, Hazen SL, Comhair SA, Erzurum SC. Oxidačné a nitrozatívne udalosti pri astme. Free Radic Biol Med. 2003;35:213�225.
17. Comhair SA, Ricci KS, Arroliga M, Lara AR, Dweik RA, et al. Korelácia systémového deficitu superoxiddismutázy s obštrukciou prietoku vzduchu pri astme. Am J Respir Crit Care Med. 2005;172:306�313.
18. Comhair SA, Xu W, Ghosh S, Thunnissen FB, Almasan A, a kol. Inaktivácia superoxiddismutázy v patofyziológii prestavby a reaktivity astmatických dýchacích ciest. Am J Pathol. 2005;166:663�674.
19. Dut R, Dizdar EA, Birben E, Sackesen C, Soyer OU, Besler T, Kalayci O. Oxidačný stres a jeho determinanty v dýchacích cestách detí s astmou. Alergia. 2008;63:1605�1609.
20. Ercan H, Birben E, Dizdar EA, Keskin O, Karaaslan C, a kol. Oxidačný stres a genetické a epidemiologické determinanty oxidačného poškodenia u detskej astmy. J Allergy Clin Immunol. 2006;118:1097�1104.
21. Fitzpatrick AM, Teague WG, Holguin F, Yeh M, Brown LA. Program výskumu ťažkej astmy. Homeostáza glutatiónu v dýchacích cestách je u detí s ťažkou astmou zmenená: dôkaz oxidačného stresu. J Allergy Clin Immunol. 2009;123:146�152.
22. Miller DM, Buettner GR, Aust SD. Prechodné kovy ako katalyzátory „autooxidačných“ reakcií. Free Radic Biol Med. 1990;8:95�108.
23. Dupuy C, Virion A, Ohayon R, Kaniewski J, D�me D, Pommier J. Mechanizmus tvorby peroxidu vodíka katalyzovaný NADPH oxidázou v plazmatickej membráne štítnej žľazy. J Biol Chem. 1991;266:3739�3743.
24. Grangerová DN. Úloha xantínoxidázy a granulocytov pri ischemiareperfúznom poškodení. Am J Physiol. 1988;255:H1269�H1275.
25. Fenton HJH. Oxidácia kyseliny vínnej v prítomnosti železa. J Chem Soc. 1984;65:899�910.
26. Haber F, Weiss JJ. Katalytický rozklad peroxidu vodíka soľami železa. Proc R Soc Lond Ser A. 1934;147:332�351.
27. Liochev SI, Fridovič I. Haber�Weiss na bicykli o 70 rokov neskôr: alternatívny pohľad. Redox Rep. 2002;7:55�57.
28. Klebanoff SJ. Myeloperoxidáza: priateľ a nepriateľ. J Leukoc Biol. 2005;77:598�625.
29. Whiteman M, Jenner A, Halliwell B. Modifikácie báz indukované kyselinou chlórnou v izolovanej DNA teľacieho týmusu. Chem Res Toxicol. 1997;10:1240�1246.
30. Kulcharyk PA, Heinecke JW. Kyselina chlórna produkovaná myeloperoxidázovým systémom ľudských fagocytov indukuje kovalentné krížové väzby medzi DNA a proteínom. Biochémia. 2001;40:3648�3656.
31. Brennan ML, Wu W, Fu X, Shen Z, Song W a kol. Príbeh dvoch kontroverzií: definovanie úlohy peroxidáz pri tvorbe nitrotyrozínu in vivo pomocou eozinofilnej peroxidázy a myší s deficitom myeloperoxidázy a povahy reaktívnych druhov dusíka generovaných peroxidázou. J Biol Chem. 2002;277:17415�17427.
32. Denzler KL, Borchers MT, Crosby JR, Cieslewicz G, Hines EM a kol. Rozsiahla degranulácia eozinofilov a peroxidázou sprostredkovaná oxidácia proteínov dýchacích ciest sa nevyskytujú v myšom modeli pľúcneho zápalu s výzvou ovalbumínu. J Immunol. 2001;167:1672�1682.
33. van Dalen CJ, Winterbourn CC, Senthilmohan R, Kettle AJ. Dusitany ako substrát a inhibítor myeloperoxidázy. Dôsledky pre nitráciu a produkciu kyseliny chlórnej v miestach zápalu. J Biol Chem. 2000;275:11638�11644.
34. Drevo LG, Fitzgerald DA, Gibson PG, Cooper DM, Garg ML. Peroxidácia lipidov stanovená izoprostánmi v plazme súvisí so závažnosťou ochorenia pri miernej astme. Lipidy. 2000;35:967�974.
35. Montuschi P, Corradi M, Ciabattoni G, Nightingale J, Kharitonov SA, Barnes PJ. Zvýšený 8-izoprostán, marker oxidačného stresu, vo vydychovanom kondenzáte pacientov s astmou. Am J Respir Crit Care Med. 1999;160:216�220.
36. Church DF, Pryor WA. Voľná radikálová chémia cigaretového dymu a jej toxikologické dôsledky. Environmentálny zdravotný výhľad. 1985;64:111�126.
37. Hiltermann JT, Lapperre TS, van Bree L, Steerenberg PA, Brahim JJ a kol. Zápal vyvolaný ozónom hodnotený v spúte a tekutine z výplachu priedušiek od astmatikov: nový neinvazívny nástroj v epidemiologických štúdiách znečistenia ovzdušia a astmy. Free Radic Biol Med. 1999;27:1448�1454.
38. Slávik JA, Rogers DF, Barnes PJ. Účinok inhalovaného ozónu na vydychovaný oxid dusnatý, funkciu pľúc a indukovaný spút u normálnych a astmatických jedincov. Hrudník. 1999;54:1061�1069.
39. Cho AK, Sioutas C, Miguel AH, Kumagai Y, Schmitz DA a kol. Redoxná aktivita polietavých častíc na rôznych miestach v povodí Los Angeles. Environ Res. 2005;99:40�47.
40. Comhair SA, Thomassen MJ, Erzurum SC. Diferenciálna indukcia extracelulárnej glutatiónperoxidázy a syntázy oxidu dusnatého 2 v dýchacích cestách zdravých jedincov vystavených 100 % O(2) alebo cigaretovému dymu. Am J Respir Cell Mol Biol. 2000;23:350�354.
41. Matthay MA, Geiser T, Matalon S, Ischiropoulos H. Oxidantom sprostredkované poškodenie pľúc pri syndróme akútnej respiračnej tiesne. Crit Care Med. 1999;27:2028�2030.
42. Biaglow JE, Mitchell JB, Held K. Význam peroxidu a superoxidu v röntgenovej reakcii. Int J Radiat Oncol Biol Phys. 1992;22:665�669.
43. Chiu SM, Xue LY, Friedman LR, Oleinick NL. Senzibilizácia miest pripojenia jadrovej matrice na ionizujúce žiarenie sprostredkovaná iónmi medi. Biochémia. 1993;32:6214�6219.
44. Narayanan PK, Goodwin EH, Lehnert BE. Alfa častice iniciujú biologickú produkciu superoxidových aniónov a peroxidu vodíka v ľudských bunkách. Cancer Res. 1997;57:3963�3971.
45. Tuttle SW, Varnes ME, Mitchell JB, Biaglow JE. Citlivosť na chemické oxidanty a žiarenie v CHO bunkových líniách s nedostatkom aktivity oxidačného pentózového cyklu. Int J Radiat Oncol Biol Phys. 1992;22: 671�675.
46. Guo G, Yan-Sanders Y, Lyn-Cook BD, Wang T, Tamae D a kol. mangán
expresia génu sprostredkovaná superoxiddismutázou pri ožarovaní
adaptívne reakcie. Mol Cell Biol. 2003;23:2362�2378.
47. Azzam EI, de Toledo SM, Spitz DR, Little JB. Oxidačný metabolizmus
moduluje prenos signálu a tvorbu mikrojadier u okoloidúcich
bunky z normálnych ľudských fibroblastov ožiarených a-časticami. Cancer Res.
2002;62:5436�5442.
48. Leach JK, Van Tuyle G, Lin PS, Schmidt-Ullrich R, Mikkelsen RB.
Ionizujúce žiarenie indukované, na mitochondriách závislé generovanie reaktívneho
kyslík/dusík. Cancer Res. 2001;61:3894�3901.
49. Dent P, Yacoub A, Fisher PB, Hagan MP, Grant S. MAPK cesty v r.
radiačné reakcie. Onkogén. 2003;22:5885�5896.
50. Wei SJ, Botero A, Hirota K, Bradbury CM, Markovina S, et al. Thioredoxin
jadrová translokácia a interakcia s redoxným faktorom-1 aktivuje transkripčný faktor AP-1 v reakcii na ionizujúce žiarenie. Cancer Res. 2000;60:6688�6695.
51. Kadet J, Douki T, Gasparutto D, Ravanat JL. Oxidačné poškodenie DNA: tvorba, meranie a biochemické vlastnosti. Mutat Res. 2003;531:5�23.
52. Yokoya A, Cunniffe SM, O�Neill P. Vplyv hydratácie na indukciu zlomov vlákien a lézií báz vo filmoch plazmidovej DNA gamažiarením. J Am Chem Soc. 2002;124:8859�8866.
53. Janssen YM, Van Houten B, Borm PJ, Mossman BT. Reakcie buniek a tkanív na oxidačné poškodenie. Lab Invest. 1993;69:261�274.
54. Iwanaga M, Mori K, Iida T, Urata Y, Matsuo T, et al. Indukcia gama glutamylcysteínsyntetázy ionizujúcim žiarením v bunkách ľudského glioblastómu T98G závislá od jadrového faktora kappa B. Free Radic Biol Med. 1998;24:1256�1268.
55. Stohs SJ, Bagchi D. Oxidačné mechanizmy v toxicite kovových iónov. Free Radic Biol Med. 1995;18:321�336.
56. Leonard SS, Harris GK, Shi X. Kovom indukovaný oxidačný stres a prenos signálu. Free Radic Biol Med. 2004;37:1921�1942.
57. Shi H, Shi X, Liu KJ. Oxidačný mechanizmus toxicity arzénu a karcinogenéza. Mol Cell Biochem. 2004;255:67�78.
58. Pi J, Horiguchi S, Sun Y, Nikaido M, Shimojo N, Hayashi T. Potenciálny mechanizmus narušenia tvorby oxidu dusnatého spôsobeného predĺženým orálnym vystavením arzenátu u králikov. Free Radic Biol Med.2003;35:102�113.
59. Rin K, Kawaguchi K, Yamanaka K, Tezuka M, Oku N, Okada S. Zlomy DNA reťazca vyvolané kyselinou dimetylarzínovou, metabolitom anorganického arzénu, sú silne zosilnené superoxidovými aniónovými radikálmi. Biol Pharm Bull. 1995;18:45�58.
60. Waalkes MP, Liu J, Ward JM, Diwan LA. Mechanizmy, ktoré sú základom karcinogenézy arzénu: precitlivenosť myší vystavených anorganickému arzénu počas tehotenstva. Toxikológia. 2004;198:31�38.
61. Schiller CM, Fowler BA, Woods JS. Účinky arzénu na aktiváciu pyruvátdehydrogenázy. Environmentálny zdravotný výhľad. 1977;19:205�207.
62. Monterio HP, Bechara EJH, Abdalla DSP. Účasť voľných radikálov na neurologických porfýriách a otravách olovom. Mol Cell Biochem. 1991;103:73�83.
63. Tripathi RM, Raghunath R, Mahapatra S. Krvné olovo a jeho vplyv na hladiny Cd, Cu, Zn, Fe a hemoglobínu u detí. Sci Total Environ. 2001;277:161�168.
64. Nehru B, Dua R. Vplyv dietetického selénu na neurotoxicitu olova. J Environ Pathol Toxicol Oncol. 1997;16:47�50.
65. Reid TM, Feig DI, Loeb LA. Mutagenéza kovmi indukovanými kyslíkovými radikálmi. Environmentálny zdravotný výhľad. 1994;102(dodatok 3):57�61.
66. Kinnula VL, Crapo JD. Superoxiddismutázy v pľúcach a ľudských pľúcnych ochoreniach. Am J Respir Crit Care Med. 2003;167:1600�1619.
67. Kinnula VL. Produkcia a degradácia metabolitov kyslíka počas zápalových stavov v ľudských pľúcach. Curr Drug Targets Inflamm Allergy. 2005;4:465�470.
68. Zelko IN, Mariani TJ, Folz RJ. Multigénová rodina superoxiddismutázy: porovnanie štruktúr, evolúcie a expresie génov CuZn-SOD (SOD1), Mn-SOD (SOD2) a EC-SOD (SOD3). Free Radic Biol Med. 2002;33:337�349.
69. Kirkman HN, Rolfo M, Ferraris AM, Gaetani GF. Mechanizmy ochrany katalázy pomocou NADPH. Kinetika a stechiometria. J Biol Chem. 1999;274:13908�13914.
70. Floh� L. Glutatiónperoxidáza. Basic Life Sci. 1988;49:663�668.
71. Arthur JR. Glutatiónperoxidázy. Cell Mol Life Sci. 2000;57:1825�1835.
72. Chu FF, Doroshow JH, Esworthy RS. Expresia, charakterizácia a tkanivová distribúcia novej bunkovej glutatiónperoxidázy závislej od selénu, GSHPx-GI. J Biol Chem. 1993;268:2571�2576.
73. Comhair SA, Bhathena PR, Farver C, Thunnissen FB, Erzurum SC. Indukcia extracelulárnej glutatiónperoxidázy v astmatických pľúcach: dôkaz redoxnej regulácie expresie v ľudských epiteliálnych bunkách dýchacích ciest. FASEB J. 2001;15:70�78.
74. Gromer S, Urig S, Becker K. Tioredoxínový systém od vedy po kliniku. Med Res Rev. 2004;24:40-89.
75. Kinnula VL, Lehtonen S, Kaarteenaho-Wiik R, Lakari E, P��kk� P, et al. Bunková špecifická expresia peroxiredoxínov v ľudskej pľúcnej a pľúcnej sarkoidóze. Hrudník. 2002;57:157�164.
76. Dubuisson M, Vander Stricht D, Clippe A, Etienne F, Nauser T a kol. Ľudský peroxiredoxín 5 je peroxynitritreduktáza. FEBS Lett. 2004;571:161�165.
77. Holmgren A. Antioxidačná funkcia tioredoxínových a glutaredoxínových systémov. Antioxidačný redoxný signál. 2000;2:811�820.
78. Dickinson DA, Forman HJ. Glutatión v obrane a signalizácii: lekcie od malého tiolu. Ann NY Acad Sci. 2002;973:488�504.
79. Sies H. Glutatión a jeho úloha v bunkových funkciách. Free Radic Biol Med. 1999;27:916�921.
80. Ladner JE, Parsons JF, Rife CL, Gilliland GL, Armstrong RN. Paralelné evolučné dráhy pre glutatión transferázy: štruktúra a mechanizmus mitochondriálneho enzýmu triedy kappa rGSTK1-1. Biochémia. 2004;43:52�61.
81. Robinson A, Huttley GA, Booth HS, Board PG. Modelovanie a bioinformatické štúdie ľudskej glutatión transferázy triedy kappa predpovedajú novú tretiu rodinu transferáz s homológiou s prokaryotickými 2-hydroxychromen-2-karboxylát izomerázami. Biochem J. 2004;379:541�552.
82. Jakobsson PJ, Morgenstern R, Mancini J, Ford-Hutchinson A, Persson B. Spoločné štrukturálne znaky rozšírenej superrodiny proteínov spojených s membránou MAPEGda s vysoko odlišnými funkciami v metabolizme eikosanoidov a glutatiónu. Protein Sci. 1999;8:689�692.
83. Hayes JD, Pulford DJ. Rodina supergénu glutatión S-transferázy: regulácia GST a príspevok izoenzýmov k chemoprotekcii rakoviny a rezistencii voči liekom. Crit Rev Biochem Mol Biol. 1995;30:445�600.
84. Armstrong RN. Štruktúra, katalytický mechanizmus a vývoj glutatión transferáz. Chem Res Toxicol. 1997;10:2-18.
85. Hayes JD, McLellan LI. Glutatión a enzýmy závislé od glutatiónu predstavujú koordinovane regulovanú obranu proti oxidačnému stresu. Free Radic Res. 1999;31:273�300.
86. Sheehan D, Meade G, Foley VM, Dowd CA. Štruktúra, funkcia a vývoj glutatión transferáz: dôsledky pre klasifikáciu necicavčích členov starodávnej nadrodiny enzýmov. Biochem J. 2001;360:1-16.
87. Cho SG, Lee YH, Park HS, Ryoo K, Kang KW a kol. Glutatión S-transferáza Mu moduluje stresom aktivované signály potlačením kinázy 1 regulujúcej signál apoptózy. J Biol Chem. 2001;276:12749�12755.
88. Dorion S, Lambert H, Landry J. Aktivácia signálnej dráhy p38 tepelným šokom zahŕňa disociáciu glutatión S-transferázy Mu z Ask1. J Biol Chem. 2002;277:30792�30797.
89. Adler V, Yin Z, Fuchs SY, Benezra M, Rosario L a kol. Regulácia signalizácie JNK pomocou GSTp. EMBO J. 1999;18:1321�1334.
90. Manevich Y, Feinstein SI, Fisher AB. Aktivácia antioxidačného enzýmu 1-CYS peroxiredoxínu vyžaduje glutationyláciu sprostredkovanú heterodimerizáciou s pGST. Proc Natl Acad Sci USA 2004;101:3780�3785.
91. Bunker VW. Voľné radikály, antioxidanty a starnutie. Med Lab Sci. 1992;49:299�312.
92. Mezzetti A, Lapenna D, Romano F, Costantini F, Pierdomenico SD a kol. Systémový oxidačný stres a jeho vzťah s vekom a chorobou. J Am Geriatr Soc. 1996;44:823�827.
93. White E, Shannon JS, Patterson RE. Vzťah medzi vitamínom a
užívanie doplnkov vápnika a rakovina hrubého čreva. Biomarkery epidemiolu rakoviny Predch. 1997;6:769�774.
94. Masella R, Di Benedetto R, Vari R, Filesi C, Giovannini C. Nové mechanizmy prírodných antioxidačných zlúčenín v biologických systémoch: účasť glutatiónu a enzýmov súvisiacich s glutatiónom. J Nutr Biochem. 2005;16:577�586.
95. Curello S, Ceconi C, Bigoli C, Ferrari R, Albertini A, Guarnieri C. Zmeny stavu srdcového glutatiónu po ischémii a reperfúzii. Skúsenosti. 1985;41:42-43.
96. El-Agamey A, Lowe GM, McGarvey DJ, Mortensen A, Phillip DM, Truscott TG. Chémia karotenoidových radikálov a antioxidačné/prooxidačné vlastnosti. Arch Biochem Biophys. 2004;430:37�48.
97. Rice-Evans CA, Sampson J, Bramley PM, Holloway DE. Prečo očakávame, že karotenoidy budú antioxidanty in vivo? Free Radic Res. 1997;26:381�398.
98. Niles RM. Signálne dráhy v retinoidnej chemoprevencii a liečbe rakoviny. Mutat Res. 2004;555:81�96.
99. Donato LJ, Noy N. Potlačenie rastu karcinómu prsníka kyselinou retinovou: proapoptotické gény sú cieľmi pre signalizáciu receptora kyseliny retinovej a bunkovej signalizácie proteínu II viažuceho kyselinu retinovú. Cancer Res. 2005;65:8193�8199.
100. Niizuma H, Nakamura Y, Ozaki T, Nakanishi H, Ohira M a kol. Bcl-2 je kľúčový regulátor pre apoptotickú bunkovú smrť indukovanú kyselinou retinovou pri neuroblastóme. Onkogén. 2006;25:5046�5055.
101. Dalton TP, Shertzer HG, Puga A. Regulácia génovej expresie reaktívnym kyslíkom. Ann Rev Pharmacol Toxicol. 1999;39:67�101.
102. Scandalios JG. Genomické reakcie na oxidačný stres. In: Meyers RA, ed. Encyklopédia molekulárnej bunkovej biológie a molekulárnej medicíny. Vol 5. 2nd ed. Weinheim, Nemecko: Wiley-VCH; 2004: 489�512.
103. Ghosh R, Mitchell DL. Vplyv oxidačného poškodenia DNA v promótorových prvkoch na väzbu transkripčného faktora. Nucleic Acids Res. 1999;27:3213�3218.
104. Marietta C, Gulam H, Brooks PJ. Jediná 8-cyklo-50-deoxyadenozínová lézia v TATA boxe bráni väzbe TATA väzbového proteínu a silne znižuje transkripciu in vivo. Oprava DNA (Amst). 20;2002:1�967.
105. Jackson AL, Chen R, Loeb LA. Vyvolanie nestability mikrosatelitov
oxidačným poškodením DNA. Proc Natl Acad Sci US A. 1998;95:12468�12473.
106. Caldecott KW. Interakcie proteín-proteín počas opravy jednovláknového zlomu cicavčej DNA. Biochem Soc Trans. 2003;31:247�251.
107. Cooke MS, Evans MD, Dizdaroglu M, Lunec J. Oxidačné poškodenie DNA: mechanizmy, mutácie a choroby. FASEB J. 2003;17:1195�1214.
108. Jones PL, Wolffe AP. Vzťahy medzi organizáciou chromatínu a metyláciou DNA pri určovaní génovej expresie. Semin Cancer Biol. 1999;9:339�347.
109. Girotti AW. Mechanizmy peroxidácie lipidov. J Free Radic Biol Med. 1985;1:87�95.
110. Siu GM, Draper HH. Metabolizmus malónaldehydu in vivo a in vitro. Lipidy. 1982;17:349�355.
111. Esterbauer H, Koller E, Slee RG, Koster JF. Možné zapojenie produktu lipidovo-peroxidácie 4-hydroxynonenalu do tvorby fluorescenčných chromolipidov. Biochem J. 1986;239:405-409.
112. Hagihara M, Nishigaki I, Maseki M, Yagi K. Zmeny hladín peroxidu lipidov v lipoproteínových frakciách ľudského séra závislé od veku. J Gerontol. 1984;39:269�272.
113. Keller JN, Mark RJ, Bruce AJ, Blanc E, Rothstein JD, et al. 4- Hydroxynonenal, aldehydový produkt peroxidácie membránových lipidov, zhoršuje transport glutamátu a mitochondriálnu funkciu v synaptozómoch. Neuroveda. 1997;806:85�96.
114. Uchida K, Shiraishi M, Naito Y, Torii Y, Nakamura Y, Osawa T. Aktivácia stresových signálnych dráh konečným produktom peroxidácie lipidov. 4-hydroxy-2-nonenal je potenciálnym induktorom intracelulárnej produkcie peroxidu. J Biol Chem. 1999;274:2234�2242.
115. Suc I, Meilhac O, Lajoie-Mazenc I, Vandaele J, Jurgens G, Salvayre R, Negre-Salvayre A. Aktivácia receptora EGF oxidovaným LDL. FASEB J. 1998;12:665�671.
116. Tsukagoshi H, Kawata T, Shimizu Y, Ishizuka T, Dobashi K, Mori M. 4-Hydroxy-2-nonenal zvyšuje produkciu fibronektínu ľudskými pľúcnymi fibroblastami IMR-90 čiastočne prostredníctvom aktivácie extracelulárneho signálu spojeného s receptorom epidermálneho rastového faktora- regulovaná dráha kinázy p44/42. Toxicol Appl Pharmacol. 2002;184:127�135.
117. Montuschi P, Collins JV, Ciabattoni G, Lazzeri N, Corradi M, Kharitonov SA, Barnes PJ. Vydychovaný 8-izoprostán ako in vivo biomarker pľúcneho oxidačného stresu u pacientov s CHOCHP a zdravých fajčiarov. Am J Respir Crit Care Med. 2000;162:1175�1177.
118. Morrison D, Rahman I, Lannan S, MacNee W. Priepustnosť epitelu, zápal a oxidačný stres vo vzduchových priestoroch fajčiarov. Am J Respir Crit Care Med. 1999;159:473�479.
119. Nowak D, Kasielski M, Antczak A, Pietras T, Bialasiewicz P. Zvýšený obsah látok reagujúcich s kyselinou tiobarbiturovou a peroxidu vodíka v kondenzáte vydychovaného dychu u pacientov so stabilnou chronickou obštrukčnou chorobou pľúc: žiadny významný vplyv fajčenia cigariet. Respir Med. 1999;93:389�396.
120. Kelly FJ, Mudway IS. Oxidácia bielkovín na rozhraní vzduch-pľúca. Aminokyseliny. 2003;25:375�396.
121. Dean RT, Roberts CR, Jessup W. Fragmentácia extracelulárnych a intracelulárnych polypeptidov voľnými radikálmi. Prog Clin Biol Res. 1985;180:341�350.
122. Keck RG. Použitie t-butylhydroperoxidu ako sondy pre oxidáciu metionínu v proteínoch. Anal Biochem. 1996;236:56-62.
123. Davies KJ. Poškodenie a degradácia bielkovín kyslíkovými radikálmi. I. Všeobecné aspekty. J Biol Chem. 1987;262:9895�9901.
124. Stadtman ER. Oxidácia proteínov katalyzovaná kovovými iónmi: biochemický mechanizmus a biologické dôsledky. Free Radic Biol Med.
1990;9:315�325.
125. Fucci L, Oliver CN, Coon MJ, Stadtman ER. Inaktivácia kľúčových metabolických enzýmov oxidačnými reakciami so zmiešanou funkciou: možný vplyv na premenu bielkovín a starnutie. Proc Natl Acad Sci US A. 1983;80:1521�1525.
126. Stadtman ER, Moskovitz J, Levine RL. Oxidácia metionínových zvyškov bielkovín: biologické dôsledky. Antioxidačný redoxný signál. 2003;5:577�582.
127. Stadtman ER, Levine RL. Voľnými radikálmi sprostredkovaná oxidácia voľných aminokyselín a aminokyselinových zvyškov v proteínoch. Aminokyseliny. 2003;25:207�218.
128. Stadtman ER. Oxidácia bielkovín pri starnutí a chorobách súvisiacich s vekom. Ann NY Acad Sci. 2001;928:22�38.
129. Shacter E. Kvantifikácia a význam oxidácie proteínov v biologických vzorkách. Drug Metab Rev. 2000;32:307�326.
130. Poli G, Leonarduzzi G, Biasi F, Chiarpotto E. Oxidačný stres a bunková signalizácia. Curr Med Chem. 2004;11:1163�1182.
131. Neufeld G, Cohen T, Gengrinovitch S, Poltorak Z. Vaskulárny endoteliálny rastový faktor (VEGF) a jeho receptory. FASEB J. 1999;13:9-22.
132. Sundaresan M, Yu ZX, Ferrans VJ, Sulciner DJ, Gutkind JS a kol. Regulácia tvorby reaktívnych druhov kyslíka vo fibroblastoch pomocou Rac1. Biochem J. 1996; 318: 379-382.
133. Sun T, Oberley LW. Redoxná regulácia transkripčných aktivátorov. Free Radic Biol Med. 1996;21:335�348.
134. Klatt P, Molina EP, De Lacoba MG, Padilla CA, Martinez-Galesteo E, Barcena JA, Lamas S. Redoxná regulácia väzby c-Jun DNA reverzibilnou S-glutatioláciou. FASEB J. 1999;13:1481�1490.
135. Reynaert NL, Clesss K, Guala AS, Wouters EF, van der Vliet A, Janssen Heininger
YM. In situ detekcia S-glutationylovaných proteínov po derivatizácii cysteínu katalyzovanej glutaredoxínom-1. Biochim Biophys Acta. 2006;1760:380�387.
136. Reynaert NL, Wouters EF, Janssen-Heininger YM. Modulácia glutaredoxínu-1
expresia v myšom modeli alergického ochorenia dýchacích ciest. Am J Respir Cell Mol Biol. 2007;36:147�151.
137. Filomeni G, Rotilio G, Ciriolo MR. Bunková signalizácia a glutatiónový redoxný systém. Biochem Pharmacol. 2002;64:1057�1064.
138. Pande V, Ramos MJ. Molekulárne rozpoznávanie 15-deoxydelta (12,14) prostaglandínu J(2) jadrovým faktorom-kappa B a inými bunkovými proteínmi. Bioorg Med Chem Lett. 2005;15:4057�4063.
139. Perkins ND. Integrácia bunkových signálnych dráh s funkciou NF-kappaB a IKK. Nat Rev Mol Cell Biol. 2007;8:49�62.
140. Gilmore TD. Úvod do NF-kappaB: hráči, cesty, perspektívy. Onkogén. 2006;25:6680�6684.
141. Hirota K, Murata M, Sachi Y, Nakamura H, Takeuchi J, Mori K, Yodoi J. Výrazné úlohy tioredoxínu v cytoplazme a v jadre. Dvojstupňový mechanizmus redoxnej regulácie transkripčného faktora NF-kappaB. J Biol Chem. 1999;274:27891�27897.
142. Oddelenie PA. Úloha komplementu, chemokínov a regulačných cytokínov pri akútnom poškodení pľúc. Ann NY Acad Sci. 1996;796:104�112.
143. Akira S, Kishimoto A. NF-IL6 a NF-kB v regulácii cytokínového génu. Adv Immunol. 1997;65:1�46.
144. Meyer M, Schreck R, Baeuerle PA. H2O2 a antioxidanty majú opačné účinky na aktiváciu NF-kappa B a AP-1 v intaktných bunkách: AP-1 ako sekundárny faktor reagujúci na antioxidanty. EMBO J. 1993;12:2005�2015.
145. Abate C, Patel L, Rausher FJ, Curran T. Redoxná regulácia väzbovej aktivity fos a jun DNA in vitro. Veda. 1990;249:1157�1161.
146. Galter D, Mihm S, Droge W. Výrazné účinky glutatióndisulfidu na jadrové transkripčné faktory kB a aktivačný proteín-1. Eur J Biochem. 1994;221:639�648.
147. Hirota K, Matsui M, Iwata S, Nishiyama A, Mori K, Yodoi J. Transkripčná aktivita AP-1 je regulovaná priamou asociáciou medzi tioredoxínom a Ref-1. Proc Natl Acad Sci US A. 1997;94: 3633�3638.
Nástroj Find A Practitioner od IFM je najväčšou sieťou odporúčaní vo funkčnej medicíne, ktorá bola vytvorená s cieľom pomôcť pacientom nájsť praktických lekárov kdekoľvek na svete. IFM Certified Practitioners sú vo výsledkoch vyhľadávania uvedení na prvom mieste vzhľadom na ich rozsiahle vzdelanie v oblasti funkčnej medicíny