Po transekcii periférneho nervu hornej končatiny a chirurgickej oprave niektorí pacienti znovu získajú dobrú senzomotorickú funkciu, zatiaľ čo iní nie. Pochopenie periférnych a centrálnych mechanizmov, ktoré prispievajú k zotaveniu, môže uľahčiť vývoj nových terapeutických zásahov. Plasticita po transekcii periférneho nervu bola preukázaná v celej neuroaxii na zvieracích modeloch poškodenia nervov. Zmeny v mozgu, ktoré sa vyskytujú po prerezaní periférnych nervov a chirurgickej oprave u ľudí, však neboli skúmané. Ďalej nebol charakterizovaný rozsah, v akom regenerácia periférnych nervov ovplyvňuje funkčné a štrukturálne zmeny mozgu. Preto sme sa pýtali, či sú funkčné zmeny sprevádzané štrukturálnymi zmenami šedej a / alebo bielej hmoty a či tieto zmeny súvisia so zmyslovým zotavením? Na riešenie týchto kľúčových problémov sme (i) hodnotili regeneráciu periférnych nervov; (ii) meranie aktivácie mozgu pomocou funkčnej magnetickej rezonancie (signál závislý od hladiny kyslíka v krvi; BOLD) ako odpoveď na vibrotaktilný stimul; (iii) skúmal štrukturálnu plasticitu mozgu šedej a bielej hmoty; a (iv) korelované merania senzorickej obnovy so zmenami šedej hmoty u pacientov s prerezaním periférnych nervov a chirurgickými opravami. V porovnaní so zdravým kontraléznym nervom každého pacienta majú prerezané nervy narušenú nervovú vodivosť 1.5 roka po pretnutí a oprave, vedú so zníženou amplitúdou a zvýšenou latenciou. V porovnaní so zdravými kontrolami mali pacienti s transekciou periférnych nervov a chirurgickou opravou zmenenú signálnu aktivitu závislú od hladiny kyslíka v krvi v kontraléznych primárnych a sekundárnych somatosenzorických kortxoch a v súbore oblastí mozgu známych ako „task positive network“. Okrem toho bolo zistené zníženie šedej hmoty v niekoľkých oblastiach mozgu, vrátane kontraléznych primárnych a sekundárnych somatosenzorických kôr, v tých istých oblastiach, kde boli identifikované zníženia signálu závislé od hladiny kyslíka v krvi. Okrem toho riedenie šedej hmoty v post-centrálnom gyruse negatívne korelovalo s mierami senzorickej obnovy (mechanická a vibračná detekcia), čo dokazuje jasné spojenie medzi funkciou a štruktúrou. Nakoniec sme identifikovali zníženú frakčnú anizotropiu bielej hmoty v pravom ostrove v oblasti, ktorá tiež preukázala zníženú šedú hmotu. Tieto výsledky poskytujú pohľad na plasticitu mozgu a vzťahy medzi štruktúrou, funkciou a správaním po poranení nervov a majú dôležité terapeutické dôsledky.
Kľúčové slová: hrúbka kôry; fMRI; difúzne zobrazovanie tenzorov; plasticita; poranenie periférnych nervov
Skratky: BA=Brodmannova oblasť; BOLD=závisí od hladiny kyslíka v krvi; fMRI = funkčná magnetická rezonancia;
PNIr = prerezanie periférneho nervu a chirurgická oprava; S1=primárna somatosenzorická kôra; S2=sekundárny somatosenzorický kortex
Obsah
úvod
Po transekcii periférneho nervu hornej končatiny a chirurgickej oprave (PNIr) sa približne 25 % pacientov nevrátilo do práce 1.5 roka po operácii (Jaquet et al., 2001). Okrem toho približne 57 % pacientov s poranením nervov je vo veku 16 – 35 rokov (McAllister et al., 1996); teda dlhý život v invalidite a ekonomické ťažkosti môžu sprevádzať transekciu nervu hornej končatiny. Pochopenie centrálnych a periférnych dôsledkov poškodenia periférnych nervov môže uľahčiť vývoj nových terapeutických stratégií a intervenčných programov.
Nie je známe, ako mozog reaguje na PNIr u ľudí. Štúdie na zvieratách však preukázali, že plasticita v rámci somatosenzorickej kôry začína bezprostredne po prerezaní periférneho nervu a že 1 rok po kompletnom prerušení nervu a chirurgickej oprave obsahujú kortikálne mapy nejednotné, nespojité znázornenie prerezaných a priľahlých nervov (Wall a kol. ., 1986). Predpokladá sa, že mechanizmy, ktoré uľahčujú funkčnú plasticitu, zahŕňajú okamžité odmaskovanie už existujúcich výbežkov zo susedných kortikálnych a subkortikálnych úrovní a dlhodobé klíčenie axónov na viacerých úrovniach neuroaxis, vrátane primárnej somatosenzorickej kôry (S1) (Florence a Kaas , 1995; Hickmott a Steen, 2005).
Štúdie zobrazovania ľudského mozgu potvrdili zistenia zo zvieracích modelov s identifikáciou zmenených funkčných aktivačných máp MRI v dôsledku poranenia miechy, amputácie, prenosu z prsta na palec a u pacientov so syndrómom karpelového tunela (Lotze et al., 2001; Manduch a kol., 2002; Jurkiewicz a kol., 2006; Napadow a kol., 2006). Okrem toho štrukturálne štúdie MRI nedávno vizualizovali zmeny šedej a bielej hmoty po traumatických poraneniach a pri rôznych patologických stavoch, vrátane amputácie končatín a chronickej bolesti (Apkarian a kol., 2004; Draganski a kol., 2006; Davis a kol., 2008; Geha a kol., 2008; máj 2008). Predpokladá sa, že zmeny šedej hmoty súvisia so zmenami veľkosti buniek, atrofiou a/alebo stratou neurónov alebo glií, zatiaľ čo zmeny bielej hmoty sú ovplyvnené axonálnou degeneráciou a stratou myelínu (Beaulieu, 2002; máj 2008).
Účinným prístupom k vymedzeniu mechanizmov patológie a plasticity je kombinácia funkčných a štrukturálnych zobrazovacích techník šedej a bielej hmoty. Už sme predtým uviedli, že pacienti s úplným PNIr hornej končatiny si zachovali hlboké somatosenzorické deficity, ktoré pretrvávajú 41.5 roka po operácii (Taylor et al., 2008a). Na základe týchto zistení sme usúdili, že títo pacienti budú vykazovať funkčné a štrukturálne zmeny mozgu v kľúčových somatosenzorických oblastiach mozgu. Preto sme v tejto štúdii predpokladali, že pacienti s PNIr by mali: (i) znížené reakcie závislé od hladiny kyslíka v krvi (BOLD) na vibračnú stimuláciu prerezaného nervového územia v oblasti S1, ktorá predstavuje poranenú hornú končatinu a v sekundárnej somatosenzorická kôra (S2); (ii) zodpovedajúce zníženie hrúbky kôry v týchto oblastiach S1 a S2 kontralézie; iii) korelácia medzi zmenami v hrúbke kôry a psychofyzickými mierami somatosenzorickej funkcie (prahové hodnoty detekcie vibrácií a dotyku); a (iv) znížená frakčná anizotropia (miera integrity bielej hmoty) v napájaní bielej hmoty do/z týchto somatosenzorických kortikálnych oblastí.
Metódy
Predmety
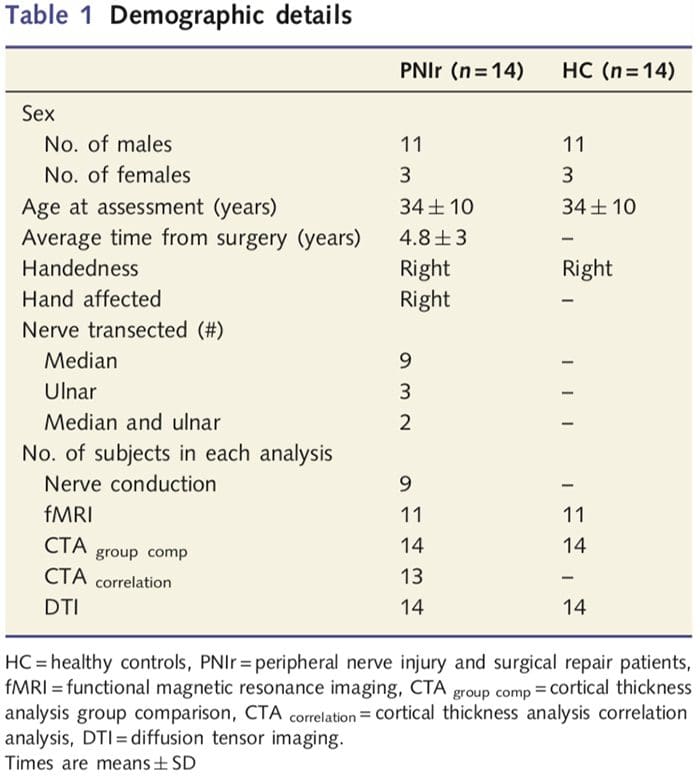
Prijali sme 27 pacientov s kompletnou transekciou stredného a/alebo lakťového nervu, po ktorej nasledovala chirurgická oprava od plastických chirurgov pridružených k University of Toronto Hand Program v období od júna 2006 do mája 2008. Z tejto väčšej kohorty bolo 14 pacientov bez bolesti (tri ženy do štúdie bolo zahrnutých 11 mužov; 34 - 10 rokov) s úplným prerezaním pravého stredného a/alebo lakťového nervu [aby sa predišlo nejasnostiam súvisiacim s prítomnosťou bolesti a laterality, pacienti s bolesťou (n=6) a ľavostrannými léziami (n=7) boli z tejto analýzy vylúčení]. Všetci pacienti podstúpili mikrochirurgickú opravu nervu najmenej 1.5 roka pred zaradením do štúdie (doba zotavenia sa pohybovala od 1.5 do 8 rokov). Okrem toho sme prijali 14 zdravých kontrol zodpovedajúcich veku a pohlavia (3 ženy, 11 mužov; 34 - 10 rokov). Všetky subjekty poskytli informovaný písomný súhlas s postupmi schválenými Etickou radou pre výskum siete univerzitného zdravia. Všetci jedinci boli praváci (určené pomocou Edinburghského inventára ruky: Oldfield, 1971) a nemali v anamnéze žiadne neurologické poškodenie alebo chronickú bolesť (či už pred alebo po pretrhnutí nervu). Demografické podrobnosti nájdete v tabuľke 1.
Študovať dizajn
Všetky subjekty sa zúčastnili na zobrazovacej relácii, ktorá zahŕňala: (i) funkčnú magnetickú rezonanciu (fMRI) ako odpoveď na vibrotaktilné stimuly aplikované na pravý ukazovák (v oblasti stredného nervu); (ii) anatomický sken celého mozgu s vysokým rozlíšením získaný na registráciu obrazu a na analýzu kortikálnej šedej hmoty; a (iii) dva difúzne tenzorové zobrazovacie skeny na posúdenie integrity bielej hmoty. Pred zobrazovaním boli subjekty poučené o základnom dizajne experimentu a pripomenuté, aby zostali čo najtichšie počas trvania skenovania.
Subjekty mohli kedykoľvek odstúpiť zo štúdie. Okrem toho sa u všetkých subjektov vykonalo senzorické a motorické hodnotenie (Taylor et al., 2008a). Keďže prahové hodnoty detekcie dotyku a vibrácií korelovali s hrúbkou kôry, opis týchto metód je uvedený nižšie (iné psychofyzikálne opatrenia budú uvedené inde).
Prah vibrácií
Prahové hodnoty detekcie vibrácií boli stanovené pomocou ručného bioteziometra (Bio-Medical Instrument Company, USA). Zariadenie má 12 mm sondu, ktorá bola umiestnená na distálnej falange pravého ukazováka (D2). Prahové hodnoty boli stanovené pomocou metódy limitov: amplitúda (napätie) sa postupne zvyšovala, až kým subjekt neukázal, že vníma podnet. Vibračné prahy sa získali trikrát a vypočítala sa priemerná hodnota. Počas testovania prahu vibrácií boli subjekty poučené, aby zavreli oči a opreli si chrbát ruky o podporný vankúš.
Prah mechanickej detekcie
Prahové hodnoty mechanickej detekcie boli stanovené pomocou štandardizovanej sady von Freyových vlákien (OptiHair2 Marstock Nervtest, Nemecko) obsahujúcej 12 logaritmicky rozmiestnených kalibrovaných vlákien, ktoré dodávali sily od 0.25 do 512 mN. Priemer kontaktnej plochy všetkých 12 vlákien bol ~ 0.4 mm. Pokusy sa uskutočnili so zatvorenými očami subjektu a rukami položenými na mäkkom vankúši. Sondy sa aplikovali vo vzostupných sériách a od subjektov sa požadovalo, aby reagovali zakaždým, keď pocítili, že sa sonda dotkla pravého konca prsta D2. Tento proces sa opakoval trikrát. Sila pre vlákno, ktorá bola zistená v najmenej dvoch z troch pokusov, bola hlásená ako prah mechanickej detekcie daného subjektu.
Testovanie nervového vedenia
Pacienti sa zúčastnili na bilaterálnych štúdiách vedenia senzorických a motorických nervov na elektromyografickej (EMG) klinike Toronto Western Hospital. Na vedenie motorického nervu sa stimulačná elektróda umiestnila na zápästie a lakeť (oddelene) a záznamová elektróda sa umiestnila na abductor pollicis brevis na hodnotenie stredného nervu alebo na abductor digiti minimi na hodnotenie ulnárneho nervu. Na testovanie senzorických nervov sa záznamová elektróda umiestnila na zápästie a stimulačná elektróda sa umiestnila na číslice D2, D3 a D5. Starší skúsený neurológ z EMG kliniky Západnej nemocnice v Toronte (Dr. Peter Ashby) preskúmal všetky klinické hodnotenia, aby určil, ktoré nervy vykazujú normálne/abnormálne reakcie. Keďže je známe, že merania amplitúdy a latencie sa medzi subjektmi značne líšia (v dôsledku faktorov, ako je hustota inervácie, hĺbka nervu a hrúbka kože jednotlivého subjektu) (Kimura, 2001), neprerušený nerv každého pacienta slúžili ako ich vlastná kontrola na porovnanie s hodnotami z prerezanej strany. U tých pacientov s detegovateľnými reakciami nervového vedenia sa vykonali párové t-testy na posúdenie rozdielu v meraniach latencie alebo amplitúdy medzi prerezanými nervami každého pacienta a netransekovanými nervami v kontralézii.
Zobrazovacie parametre
Dáta zo zobrazovania mozgu sa získali pomocou systému 3T GE MRI vybaveného osemkanálovou hlavovou cievkou s fázovým poľom. Subjekty boli umiestnené na chrbte na MRI stôl a hlava každého subjektu bola podložená, aby sa znížil pohyb. Údaje fMRI z celého mozgu boli získané pomocou echo planárneho zobrazovania (28 axiálnych rezov, zorné pole (FOV) = 20 x 20 cm, 64 x 64 matrica, 3.125 x 3.125 x 4 mm voxely, čas ozveny (TE) = 30 ms, opakovanie čas (TR) = 2000 ms). Čas skenovania bol 5 minút a 8 s (154 snímok). Počas skenovania bol aplikovaný nebolestivý, 12 Hz vibrotaktilný stimul na distálnu falangu pravej D2 pomocou balónových membrán poháňaných stlačeným vzduchom (zariadenie vyrobené Dr Christo Pantev; www.biomag.uni-muenster.de). Stimuly boli dodávané v blokoch 10 s preložených 20 s prestávkami, celkovo 10 blokov stimulácie a 10 blokov odpočinku. Prvých 8 s (4 TR) údajov získaných z každého cyklu bolo vyradených, aby sa umožnila ekvilibrácia signálu fMRI. Subjekty boli poučené, aby mali počas skenovania zatvorené oči a sústredili sa na podnety. Trojrozmerný (3D) anatomický sken celého mozgu s vysokým rozlíšením (124 sagitálnych rezov, 24 x 24 cm FOV, 256 x 256 matrica, 1.5 x 0.94 x 0.94 mm voxelov) sa získal pomocou T1-váženej 3D rozmaznanej gradientovej echo sekvencie (priemer jedného signálu, uhol preklopenia = 20°, TE < 5 ms). Okrem toho sa získali dva difúzne tenzorové zobrazovacie skeny (38 axiálnych rezov, FOV 24 x 24 cm, 128 x 128 matrica, 1.875 x 1.875 x 3 mm voxely) v 23 smeroch s hodnotou b 1000smm�2. Každý cyklus tiež obsahoval dva objemy bez difúzneho váženia.
Analýza fMRI
Dáta boli analyzované pomocou Brainvoyager QX v1.8 (Brain Innovaton, Maastricht, Holandsko). Predspracovanie zahŕňalo: korekciu 3D pohybu, korekciu času skenovania rezu, lineárne odstraňovanie trendov, filtrovanie horných priepustov (päť cyklov na beh) a priestorové vyhladzovanie s 6 mm plnou šírkou pri polovičnom maxime (FWHM) Gaussovho jadra. Súbory údajov fMRI boli interpolované na 3 x 3 x 3 mm voxely, zaregistrované do anatomického obrazu s vysokým rozlíšením a normalizované na štandardný Talairachov priestor (Talairach a Tournoux, 1988). Voxely sú uvádzané ako 1 x 1 x 1 mm. Údaje sa analyzovali použitím všeobecného lineárneho modelu; model bol získaný konvolúciou funkcie boxcar časového priebehu taktilnej stimulácie so štandardnou funkciou hemodynamickej odozvy. Na identifikáciu rozdielov medzi skupinami v aktivačných vzorcoch sa uskutočnila analýza fixných účinkov s kontrastmi: (i) zdravé kontroly: stimulácia 4 odpočinok; (ii) PNIr: stimulácia 4 odpočinok; a (iii) zdravé kontroly 4 PNIr. Aktivačné mapy boli limitované na opravenú hodnotu P50.05 (odvodené z nekorigovaných P50.0001 a 120 mm3 súvislých voxelov, ako už bolo uvedené: Taylor a Davis, 2009); toto bolo tiež overené spustením simulácie Monte Carlo s aplikáciou AlphaSim implementovanou v softvéri analýzy funkčného neuroobrazu (AFNI). Táto analýza zahŕňala iba 11 pacientov, u ktorých pretrvávala transekcia pravého stredného nervu (n = 9) alebo pravého stredného a lakťového nervu (n = 2) (tj traja pacienti s čistou transekciou pravého lakťového nervu neboli zahrnutí do tejto analýzy ).
Analýza kortikálnej hrúbky
Analýza hrúbky kortikálnej kôry sa uskutočnila pomocou Freesurfer (http://surfer.nmr.mgh.harvard.edu); metódy boli podrobne opísané inde (Dale a kol., 1999; Fischl a kol., 1999a, b; Fischl a Dale 2000). Stručne povedané, súbory anatomických údajov s vysokým rozlíšením T1 boli zaregistrované v atlase Talairach (Talairach a Tournoux, 1988). Nasledovala normalizácia intenzity, odizolovanie lebky a oddelenie hemisfér. Následne boli identifikované a segmentované hranice biela/sivá hmota (nazývaná biely povrch) a sivá/CSF (nazývaná povrch pial). Vzdialenosť medzi bielym a pialovým povrchom bola potom vypočítaná v každom bode v každej hemisfére mozgu. Aby sa identifikovali skupinové rozdiely medzi 14 pacientmi a 14 vekovo/pohlavie zodpovedajúcimi kontrolami, vykonala sa všeobecná lineárna modelová analýza v každom bode mozgu. Keďže kortikálna topografia jednotlivca je vo svojej podstate heterogénna, pred štatistickou analýzou sa použilo 5 mm priestorové vyhladzovacie jadro FWHM. Dáta sú zobrazené s opraveným P50.05 (odvodeným z nekorigovaného P50.0075 a 102 susediacich vrcholov); to bolo vypočítané spustením simulácie Monte Carlo s AlphaSim. Vrchol predstavuje bod na dvojrozmernom liste a v tejto štúdii je vzdialenosť medzi dvoma vrcholmi 0.80 mm2.
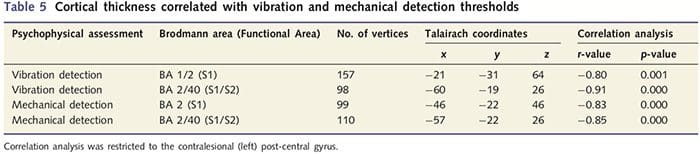
Keďže pacienti vykazovali významné deficity v somatosenzorickej funkcii v preseknutom nervovom území, predpokladali sme, že merania somatosenzorickej funkcie (detekcia vibrácií a dotyku) by korelovali s hrúbkou kôry v kontraléznom postcentrálnom gyre (primárne a sekundárne somatosenzorické kortexy). Preto sme v skupine pacientov vykonali korelačné analýzy medzi: (i) hrúbkou kôry a prahom detekcie vibrácií; a (ii) prahy detekcie hrúbky kôry a dotyku. Jeden pacient nedokončil psychofyzikálne vyšetrenie; preto táto analýza zahŕňala 13 pacientov s PNIr. Okrem toho, aby sa určilo, či existuje vzťah medzi hrúbkou kôry a časom zotavenia, bola medzi týmito dvoma meraniami vykonaná aj korelačná analýza. Tieto korelačné analýzy boli obmedzené na kontralézny post-centrálny gyrus zahrnutím masky (prevzatej zo zabudovaného atlasu Freesurferov) do všeobecného lineárneho modelu. Uskutočnila sa simulácia Monte Carlo, ktorá bola obmedzená na počet vrcholov v rámci post-centrálneho gyru kontralézie; obrázky sú zobrazené s opraveným P50.05 (odvodeným z neopraveného P50.0075 a 68 susediacich vrcholov).
Analýza zobrazovania difúzneho tenzora
Spracovanie obrazu difúzneho tenzora sa uskutočnilo pomocou DTiStudio (www.MriStudio.org) a FSLv.4.0 (www.fmrib.ox.ac.uk/fsl/). Snímky boli najprv prerovnané pomocou nástroja na automatickú registráciu obrázkov implementovaného v DTiStudio, pričom sa ako šablóna použil prvý obrázok B0 v prvej sérii. Tento proces koriguje pohyb objektu a skreslenie vírivými prúdmi. Všetky obrázky sa potom vizuálne skontrolovali, aby sa posúdila kvalita obrazu a zarovnanie samostatných zobrazovacích cyklov difúzneho tenzora. Ak sa detegoval artefakt, pred výpočtom priemeru dvoch samostatných cyklov zobrazovania difúzneho tenzora sa rez odstránil. Jednotlivé mapy FA boli vypočítané pomocou nástroja DTIFIT implementovaného vo FSL. Uskutočnila sa voxelová štatistická analýza na identifikáciu skupinových rozdielov v priemernej frakčnej anizotropii s použitím priestorovej štatistiky založenej na traktoch; pre úplný opis týchto metód pozri Smith et al. (2006). Stručne povedané, obrázky boli nelineárne zaregistrované na cieľový obrázok (MNI152), stredný obrázok sa potom vytvoril zo všetkých súborov údajov a tento obrázok sa následne stenčil, aby reprezentoval všetky trakty, ktoré boli spoločné pre všetky subjekty. Najvyššie hodnoty frakčnej anizotropie každého subjektu sa potom premietli na kostru hľadaním v bielej hmote kolmo na každý bod na kostre bielej hmoty. Potom sa medzi skupinami (14PNIr a 14 zdravých kontrol) uskutočnila štatistická analýza celého mozgu voxelom a snímky sa korigovali na celý mozog pri P50.05. Okrem toho sa v bielej hmote uskutočnila analýza oblasti záujmu traktoch susediace s kontralaterálnou S1, talamom a bilaterálnou prednou a zadnou inzulou. Tieto oblasti boli vybrané, pretože sa predtým podieľali na aspektoch somatosenzácie a pretože zodpovedajú oblastiam, ktoré boli identifikované v skupinových analýzach fMRI a analýzy kortikálnej hrúbky (CTA). Oblasti záujmu sa nakreslili na kostru bielej hmoty nasledovne: (i) Kontralaterálna oblasť záujmu S1 vznikla mediálne na spojnici medzi kostrou bielej hmoty corona-radiata a sekciou kostry napájajúcou sa do postcentrálneho gyru; končiace na konci traktu v rámci daného rezu.
V smere z oblasť záujmu siahala od z=49 do 57; trakty bielej hmoty zásobujúce oblasť rúk. (ii) Kontralaterálna oblasť záujmu talamu bola obmedzená na stopy bielej hmoty obklopujúce zadné a stredné talamické jadrá (jadrá zapojené do somatosenzorickej funkcie), siahajúce od z = �1 do 4. (iii) Ostrovné oblasti záujmu boli nakreslené bilaterálne v rámci biela hmota susediaca s prednou a zadnou izoláciou na základe kritérií, ktoré predtým zverejnilo naše laboratórium (Taylor et al., 2008b). Oblasť záujmu siahala od z = 2 do 8. Hodnoty frakčnej anizotropie sa extrahovali z každej z týchto oblastí záujmu a vykonala sa multivariačná analýza rozptylu (MANOVA) s použitím štatistického balíka pre sociálne vedy v13.0 (SPSS Inc, Chicago), ktorý zahŕňal hodnoty frakčnej anizotropie pre všetkých šesť oblastí záujmu.
výsledky
Tabuľka 1 poskytuje demografické podrobnosti pre účastníkov štúdie. Všetkých 14 pacientov prekonalo kompletnú transekciu pravého stredného a/alebo ulnárneho nervu, po ktorej nasledovala mikrochirurgická oprava aspoň 1.5 roka pred zaradením do štúdie. Čas od operácie po testovanie sa pohyboval od 1.5 do 8 rokov s priemerom (?SD) 4.8 ? 3 roky. Pacienti a kontroly nie (34 - 10 rokov obe skupiny; t = 0.04; P = 0.97).
Psychofyzika
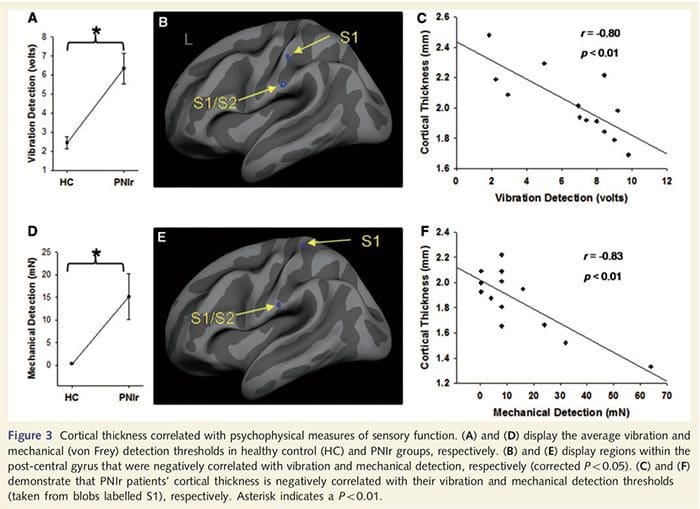
Vibračné prahy boli vypočítané zo všetkých troch meraní, pretože jednosmerná analýza rozptylu opakovaných meraní (ANOVA) neukázala žiadne významné rozdiely medzi tromi pokusmi [F (25, 1) = 0.227, P = 0.64]. Prahové hodnoty vibrácií a mechanickej detekcie boli významne narušené u pacientov s PNIr v porovnaní so zdravými kontrolami (vibrácie: t = 4.77, P50.001, obr. 3A; mechanické: t=3.10, P=0.005, obr. 3D).
Testovanie nervového vedenia
Merania amplitúdy a latencie získané z kontraléznych nervov každého pacienta boli skúseným neurológom na EMG klinike Západnej nemocnice v Toronte klasifikované ako normálne. Deväť zo 14 pacientov dokončilo test nervového vedenia. Tabuľka 2 zobrazuje priemerné zvýšenie/zníženie latencie a amplitúdy pre senzorické vedenie nervov zo zápästia do abductor pollicis brevis (stredná) alebo abduktora digiti minimi (ulnárne) a pre zmyslové vedenie zo zápästia do D2 (stredná) a D5 ( ulnar) v porovnaní s každým pacientom neporanený protilezný nerv. Z deviatich malo sedem pacientov transekcie, ktoré zahŕňali stredný nerv. Z týchto siedmich pacientov nemal jeden pacient žiadnu zistiteľnú odpoveď počas testovania motoriky a ďalší pacient nemal žiadnu detekovateľnú odpoveď počas senzorického testovania.
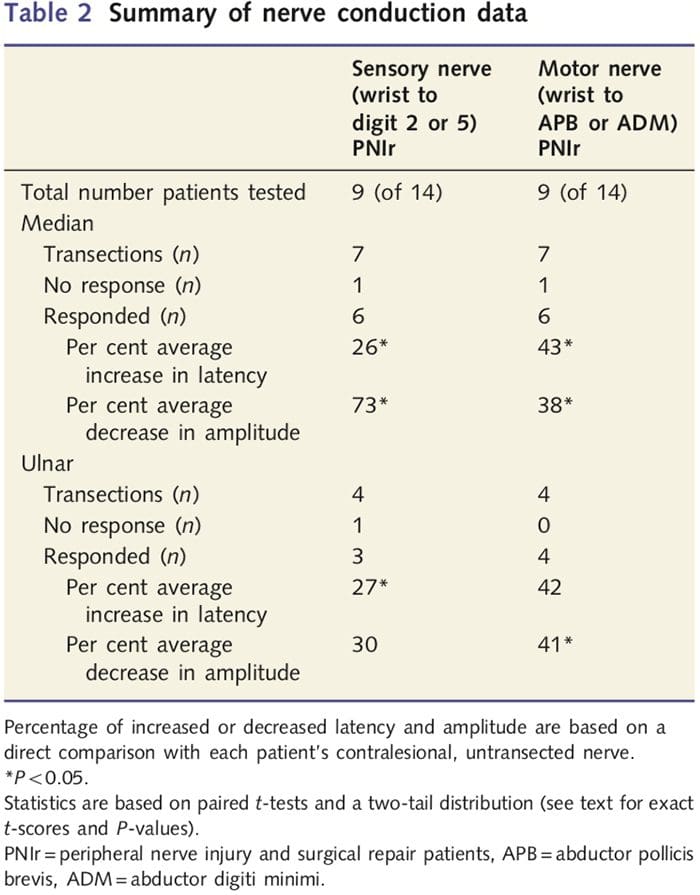 U šiestich pacientov s detegovateľnými odpoveďami sa latencie motorického vedenia zvýšili o 43 % (t=6.2; P=0.002) a amplitúdy sa znížili o 38 % (t=�2.6; P=0.045), keď bol prerezaný nerv každého pacienta v porovnaní s ich nezranenou stranou. Senzorické vedenie v stredných nervoch tiež odhalilo 26% zvýšenie latencie (t=3.9; P=0.011) a 73% zníženie amplitúdy (t=�8.0; P=0.000) v porovnaní s normálnymi kontraléznymi nervami. U štyroch pacientov s prerezaním ulnárneho nervu nemal jeden pacient žiadnu detegovateľnú odpoveď počas testovania senzorického nervu. U týchto pacientov s odpoveďami neboli motorické latencie ulnárneho nervu významne zvýšené (t = 2.8; P = 0.070); amplitúdy však boli významné
U šiestich pacientov s detegovateľnými odpoveďami sa latencie motorického vedenia zvýšili o 43 % (t=6.2; P=0.002) a amplitúdy sa znížili o 38 % (t=�2.6; P=0.045), keď bol prerezaný nerv každého pacienta v porovnaní s ich nezranenou stranou. Senzorické vedenie v stredných nervoch tiež odhalilo 26% zvýšenie latencie (t=3.9; P=0.011) a 73% zníženie amplitúdy (t=�8.0; P=0.000) v porovnaní s normálnymi kontraléznymi nervami. U štyroch pacientov s prerezaním ulnárneho nervu nemal jeden pacient žiadnu detegovateľnú odpoveď počas testovania senzorického nervu. U týchto pacientov s odpoveďami neboli motorické latencie ulnárneho nervu významne zvýšené (t = 2.8; P = 0.070); amplitúdy však boli významné
znížil o 41 % (t = �5.9; P = 0.010). Senzorické testovanie ulnárneho nervu preukázalo 27 % zvýšenie latencie (t = 4.3; P = 0.049), ale žiadne významné zvýšenie amplitúdy (t = 3.5; P = 0.072).
Funkčná plasticita v primárnom somatosenzorickom kortexe
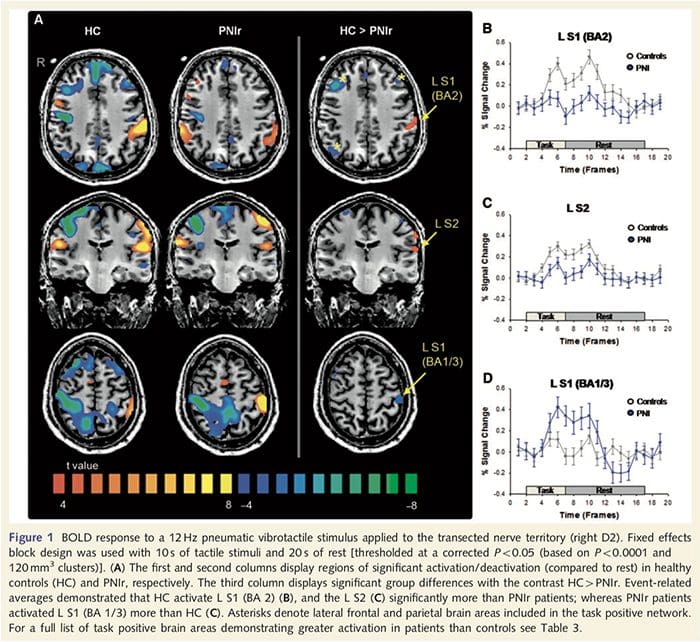
Funkčné mapy MRI sa vypočítali z 11 pacientov s PNIr s prerezaním pravého stredného nervu (pacienti s prerezaním ulnárneho nervu boli z tejto analýzy vylúčení) a z 11 zdravých kontrol zodpovedajúcich veku a pohlavia. Z obr. 1A je zrejmé, že pacienti s PNIr majú v porovnaní so zdravými kontrolami výrazne menšiu aktiváciu v oblasti S1 zodpovedajúcej oblasti Brodmann 2 (BA2) (Talairach a Tournoux, 1988) a S2 (podrobnosti nájdete v tabuľke 3). . Priemerné reakcie súvisiace s udalosťou z týchto oblastí záujmu zdôrazňujú oslabenú odpoveď BOLD v rámci pacientov vľavo BA2 a vľavo S2 (obr. 1B a C, v tomto poradí). Je zvláštne, že vibrotaktilná stimulácia u pacientov aktivovala lepšiu časť postcentrálneho gyru (pravdepodobne BA1/3) (Talairach a Tournoux, 1988) (obr. 1A a tabuľka 3). Priemer súvisiaci s udalosťou (obr. 1D) ukazuje, že zdravé kontroly mali v tejto oblasti minimálnu aktiváciu. Okrem toho pacienti mali výrazne väčšiu aktiváciu v oblastiach mozgu, ktoré sú súhrnne známe ako sieť s pozitívnymi úlohami (hviezdičky na obr. 1). Úplný zoznam aktivovaných oblastí mozgu s pozitívnymi úlohami nájdete v tabuľke 3. Táto sieť zahŕňa laterálne prefrontálne, laterálne parietálne, premotorické a inferiorné temporálne kôry (tabuľka 3): mozgové oblasti, ktoré sú aktivované počas vykonávania úlohy vyžadujúcej pozornosť a potlačené alebo neaktívne počas odpočinku alebo úloh, ktoré nie sú náročné na kognitívne alebo pozornosť (Fox et al., 2005; DeLuca a kol., 2006; Seminowicz a Davis 2007).
Znížená sivá hmota v primárnej somatosenzorickej kôre koreluje s regeneráciou zmyslov
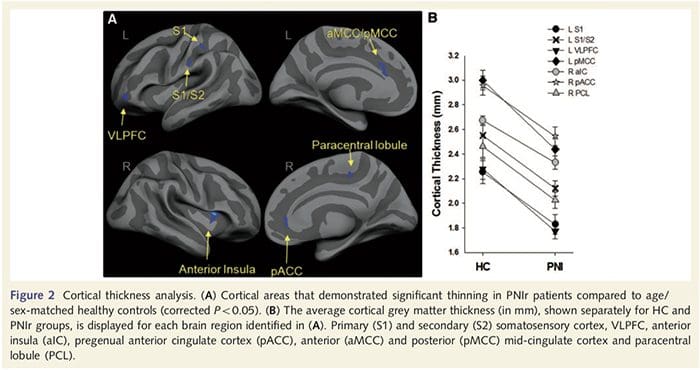
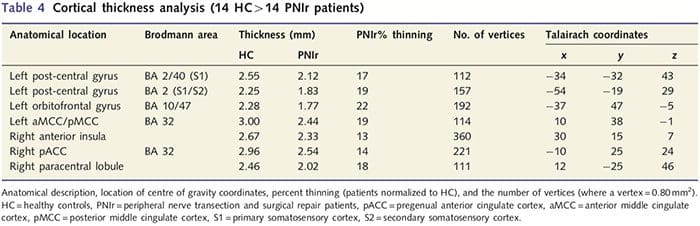
Analýza hrúbky kôry u všetkých 14 pacientov a 14 zdravých kontrol s vekom/pohlavím odhalila niekoľko lokusov významného stenčenia kôry v skupine PNIr (obr. 2 a tabuľka 4). Konkrétne, pacienti mali 13% – 22% redukciu hrúbky kôry v ľavom (kontralezionálnom) S1, S2, pregennom prednom cingulárnom gyre, ventrolaterálnom prefrontálnom kortexe a pravej prednej insule, prednom/zadnom strednom cingulárnom gyre a paracentrálnom laloku. Je zaujímavé, že miesta stenčovania šedej hmoty v post-centrálnom gyre sa zhodujú s oblasťami zníženého BOLD po vibrotaktilnej stimulácii (tabuľka 4). Keďže sme mali predchádzajúce znalosti o senzorických deficitoch a čase zotavenia pacientov (tj čas od mikrochirurgickej opravy), ďalej sme sa pýtali, či hrúbka kortikálnej kôry pacientov v postcentrálnom gyrus koreluje s ich senzorickými prahmi detekcie mechanických a vibrácií, alebo s ich čas obnovenia. Tieto analýzy odhalili negatívnu koreláciu medzi hrúbkou kôry a prahovými hodnotami detekcie vibrácií v oblasti zahŕňajúcej BA1/2 a S2 (P50.001, r = ≥ 0.80 a ≥ 0.91, pre BA1/2 a S2, v tomto poradí; obr. 3 a tabuľka 5 ). Okrem toho prahy mechanickej detekcie tiež negatívne korelovali s hrúbkou kôry v mierne lepšej oblasti BA2 a rovnakej oblasti S2 (P50.001, r = ≤ 0.83 a ≤ 0.85 pre BA2 a S2, v tomto poradí; obr. 3 a tabuľka 5). Neidentifikovali sme však významný vzťah medzi časom zotavenia a hrúbkou kôry. Preto v postcentrálnom gyruse kortikálne stenčenie bolo spojené so závažnejšími senzorickými deficitmi. Neidentifikovali sme však významný vzťah medzi časom zotavenia a hrúbkou kôry. Opäť existovala zhoda medzi kortikálnym stenčovaním v oblastiach negatívne korelujúcich s vibračnými stimulmi a oblasťami vykazujúcimi skupinové abnormality fMRI a CTA.
Abnormality bielej hmoty po transekcii nervu
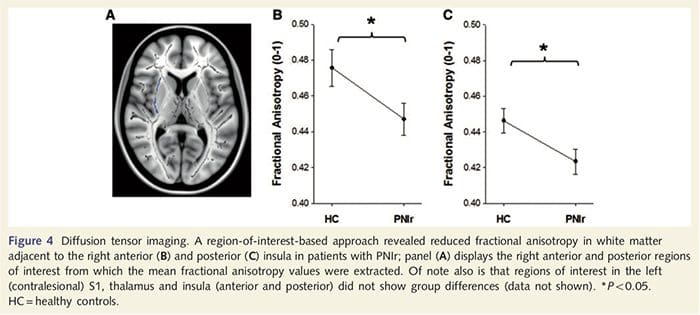
Na posúdenie integrity bielej hmoty sme použili prístup v oblasti záujmu na preskúmanie rozdielov medzi skupinami bielej hmoty na základe apriórnych hypotéz. Oblasti záujmu boli obmedzené na úseky bielej hmoty obklopujúce a napájajúce sa do kontralézie S1 a talamu. Okrem toho sme tiež nakreslili oblasti záujmu v bielej hmote susediace s ľavým a pravým, predným a zadným ostrovom. Ostrovček bol vybraný, pretože sa podieľa na somatosenzorickom spracovaní a pretože sme identifikovali zníženú šedú hmotu v pravom prednom ostrovčeku s CTA. Tento prístup v oblasti záujmu odhalil, že pacienti mali signifikantne znížené hodnoty frakčnej anizotropie bielej hmoty (MANOVA vrátane všetkých šiestich oblastí záujmu) v blízkosti pravej prednej časti [F (1, 26) = 4.39, P = 0.046; Obr. 4A] a zadná insula [F (1, 26) = 5.55, P = 0.026; Obr. 4B], ale nezistili sa žiadne skupinové rozdiely v bielej hmote susediacej s ľavým ostrovčekom (ľavý predný ostrovček: P = 0.51; ľavý zadný ostrovček: P=0.26), talame (P=0.46) alebo S1 (P=0.46 ).
 Diskusia
Diskusia
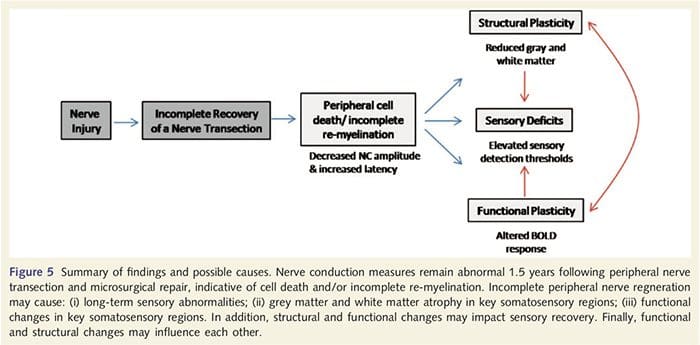
Tu sme prvýkrát preukázali, že po transekcii periférneho nervu hornej končatiny a chirurgickej oprave existuje funkčná plasticita a štrukturálne abnormality šedej aj bielej hmoty v niekoľkých kortikálnych oblastiach. Táto plasticita môže vzniknúť z neúplnej regenerácie periférnych nervov (periférna bunková smrť a/alebo neúplná re-myelinizácia), keďže merania nervového vedenia u týchto pacientov preukázali závažné abnormality. Naše údaje navyše ukazujú, že znížené vibrotaktilne vyvolané fMRI reakcie v post-centrálnom gyruse korešpondujú s riedením šedej hmoty v skupine pacientov. Tieto výsledky naznačujú, že znížené reakcie BOLD môžu byť uľahčené znížením kortikálnej šedej hmoty a / alebo znížením aferentného vstupu do post-centrálneho gyru. Okrem toho hrúbka kôry v tých istých častiach post-centrálneho gyru negatívne korelovala s behaviorálnymi mierami somatosenzorickej funkcie. To znamená, že zvýšené somatosenzorické deficity korelovali s tenšou kôrou; oboje môže súvisieť s aferentným vstupom. Celkovo možno povedať, že naše údaje naznačujú, že neúplná regenerácia periférnych nervov prispieva k somatosenzorickým poruchám, atrofii kortikálnej šedej hmoty a zníženej aktivácii fMRI (súhrn týchto zistení pozri na obr. 5).
Je dobre známe, že kortikálna plasticita po pretrhnutí periférneho nervu a chirurgickej oprave sa môže vyskytnúť v celom CNS u primátov (okrem človeka) (Kaas, 1991). Predpokladá sa, že táto plasticita je spôsobená demaskovaním predtým tichých synapsií alebo axonálneho klíčenia do deaferentovaného územia (Wall a kol., 1986; Florence a Kaas, 1995). V modeli primátov, 1 rok po pretrhnutí nervu a chirurgickej oprave, je denervovaná kôra charakterizovaná neúplnými a neusporiadanými reprezentáciami regenerovaných a priľahlých (intaktných) nervov. Táto nejednotná reprezentácia sa pripisuje neúplnej periférnej regenerácii, ktorá vedie k čiastočnému zotaveniu denervovaného kortikálneho priestoru (Kaas, 1991). Aby sme zhodnotili rozsah periférnej regenerácie v našej populácii pacientov, vykonali sme štúdie vedenia senzorických a motorických nervov cez prerezanú oblasť. Naše výsledky nervového vedenia ukazujú, že pacienti s PNIr majú významne zníženú amplitúdu a zvýšenú latenciu v senzorických aj motorických nervoch v porovnaní s ich vlastnou neprerušenou stranou. Znížená amplitúda kombinovaná so zvýšenou latenciou je indikátorom straty periférnych vlákien (tj bunkovej smrti) a/alebo abnormálnej alebo neúplnej re-myelinizácie po transekcii (Kimura, 1984). Okrem toho je dobre známe, že 20 % až 50 % neurónov ganglií dorzálnych koreňov odumiera po prerušení nervu (Liss a kol., 1996). Smrť aferentných buniek a neúplná regenerácia teda môžu viesť k zníženiu aferentného vstupu do kôry, čo môže zodpovedať za prebiehajúce senzorické deficity a zníženú odpoveď BOLD v BA2 a S2. Okrem toho by tento znížený aferentný vstup mohol tiež zodpovedať za kortikálne stenčenie, ktoré sme pozorovali v rovnakých oblastiach kôry. Ukázalo sa, že senzorická deprivácia spôsobuje transneurónovú degeneráciu v niekoľkých oblastiach CNS, vrátane dorzálneho rohu po sekcii sedacieho nervu (Knyihar-Csillik et al., 1989) a môže zahŕňať neuróny druhého a tretieho rádu (Powell a Erulkar, 1962). Transneuronálna degenerácia je charakterizovaná zmršťovaním buniek a predpokladá sa, že súvisí so zníženým alebo neexistujúcim aferentným vstupom (Knyihar-Csillik et al., 1989). Strata kortikálnej šedej hmoty (alebo atrofia) by teda mohla tiež priamo súvisieť so zníženým aferentným vstupom.
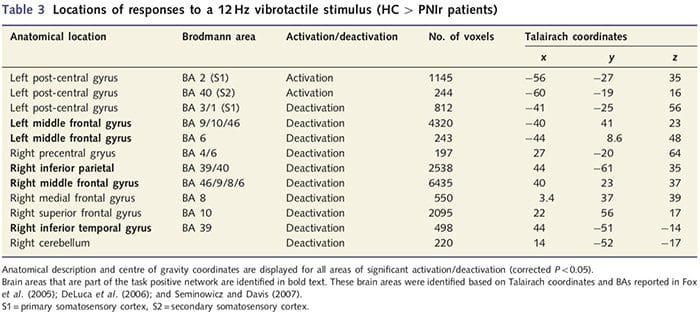
 Preukázali sme tiež zvýšenú aktiváciu v post-centrálnom gyre v oblasti zodpovedajúcej BA1/3 (Talairach a Tournoux, 1988). Elektrofyziologické, anatomické sledovacie a neuroimagingové štúdie preukázali, že pre väčšinu kožných mechanoreceptívnych aferentov sú prvými kortikálnymi destináciami BA1 a BA3b. Každá z týchto cytoarchitektonických oblastí mozgu má somatotopickú mapu tela s malými receptívnymi poľami. Okrem toho tieto oblasti reagujú na mnohé vlastnosti hmatových informácií, ako je textúra a drsnosť, rýchlosť a zakrivenie podnetov (Bodegard et al., 2001). Štúdie fMRI ukázali, že aktivita v rámci somatosenzorickej kôry je ovplyvnená pozornosťou tak, že reakcie fMRI na hmatové stimuly v S1 sú zvýšené, keď sa subjekty venujú hmatovému stimulu, ale sú oslabené, keď sú subjekty rozptýlené (Arthurs et al., 2004; Porro a kol., 2004). Okrem toho naši pacienti aktivovali sieť mozgových oblastí známych ako task positive network (DeLuca et al., 2006) viac ako zdravé kontroly. Tieto oblasti mozgu sa aktivujú počas procesov náročných na pozornosť (Fox a kol., 2005; Seminowicz a Davis, 2007). Tieto zistenia spolu naznačujú, že pacienti sa musia venovať stimulu viac ako kontrolám z dôvodu ich narušeného zmyslového vstupu. Táto zvýšená pozornosť môže tiež zodpovedať za zvýšenú aktiváciu v BA1/3b. Samozrejme, zvýšená aktivácia v BA1/3b môže odrážať aj plasticitu, ktorá nesúvisí so záťažou pozornosti.
Preukázali sme tiež zvýšenú aktiváciu v post-centrálnom gyre v oblasti zodpovedajúcej BA1/3 (Talairach a Tournoux, 1988). Elektrofyziologické, anatomické sledovacie a neuroimagingové štúdie preukázali, že pre väčšinu kožných mechanoreceptívnych aferentov sú prvými kortikálnymi destináciami BA1 a BA3b. Každá z týchto cytoarchitektonických oblastí mozgu má somatotopickú mapu tela s malými receptívnymi poľami. Okrem toho tieto oblasti reagujú na mnohé vlastnosti hmatových informácií, ako je textúra a drsnosť, rýchlosť a zakrivenie podnetov (Bodegard et al., 2001). Štúdie fMRI ukázali, že aktivita v rámci somatosenzorickej kôry je ovplyvnená pozornosťou tak, že reakcie fMRI na hmatové stimuly v S1 sú zvýšené, keď sa subjekty venujú hmatovému stimulu, ale sú oslabené, keď sú subjekty rozptýlené (Arthurs et al., 2004; Porro a kol., 2004). Okrem toho naši pacienti aktivovali sieť mozgových oblastí známych ako task positive network (DeLuca et al., 2006) viac ako zdravé kontroly. Tieto oblasti mozgu sa aktivujú počas procesov náročných na pozornosť (Fox a kol., 2005; Seminowicz a Davis, 2007). Tieto zistenia spolu naznačujú, že pacienti sa musia venovať stimulu viac ako kontrolám z dôvodu ich narušeného zmyslového vstupu. Táto zvýšená pozornosť môže tiež zodpovedať za zvýšenú aktiváciu v BA1/3b. Samozrejme, zvýšená aktivácia v BA1/3b môže odrážať aj plasticitu, ktorá nesúvisí so záťažou pozornosti.
 BA2 a S2 dostávajú projekcie z BA1/3b a tiež z odlišných častí ventroposteriorného talamického komplexu (Pons a kol., 1985; Friedman a Murray, 1986). Oba tieto mozog oblasti majú veľké, často viacmiestne (BA2) alebo bilaterálne (S2) receptívne polia (Pons et al., 1985; Iwamura et al., 2002). Na základe anatomických projekcií a vlastností neurónovej odozvy bolo preukázané hierarchické spracovanie hmatových informácií od BA1/3b po BA 2 (Kaas et al., 2002). Okrem toho elektrofyziologické štúdie na makakoch (Pons a kol., 1987) a údaje z magnetoencefalografie získané u ľudí naznačujú, že sériové spracovanie hmatových vstupov prebieha od S1 po S2 u vyšších primátov (Frot a Mauguiere 1999; Disbrow a kol. ., 2001). Niekoľko štúdií preukázalo, že BA2 je prednostne aktivovaný tvarom a zakrivením (Bodegard a kol., 2001), zatiaľ čo S2 sa môže podieľať na hmatovom učení (Ridley a Ettlinger 1976; Murray a Mishkin, 1984), čo podporuje názor, že tieto oblasti mozgu sa podieľajú na somatosenzorickom spracovaní vyššieho rádu. Naše psychofyzikálne hodnotenie preukázalo, že pacienti boli významne oslabení pri detekcii jednoduchých hmatových stimulov a v teste identifikácie textúry tvaru 1.5 roka po operácii (Taylor et al., 2008a). Tento posledný test hodnotí schopnosť pacienta rozpoznať vlastnosti objektu pri aktívnom skúmaní tvaru alebo štruktúry, čo si vyžaduje integráciu zmyslových informácií v rôznych oblastiach tela (Rosen a Lundborg, 1998). Celkovo možno povedať, že jednou z interpretácií našich údajov je, že pacienti s PNIr sa viac venujú vibrotaktilným stimulom, čo vedie k zvýšenej aktivácii siete pozitívnej úlohy a BA1/3. U týchto pacientov však naše údaje naznačujú, že oblasti spracovania vyššieho rádu, ako napríklad BA2 a S2, nedostali hmatové informácie, čo zase môže viesť k stenčeniu kôry a zníženiu BOLD odpovedí.
BA2 a S2 dostávajú projekcie z BA1/3b a tiež z odlišných častí ventroposteriorného talamického komplexu (Pons a kol., 1985; Friedman a Murray, 1986). Oba tieto mozog oblasti majú veľké, často viacmiestne (BA2) alebo bilaterálne (S2) receptívne polia (Pons et al., 1985; Iwamura et al., 2002). Na základe anatomických projekcií a vlastností neurónovej odozvy bolo preukázané hierarchické spracovanie hmatových informácií od BA1/3b po BA 2 (Kaas et al., 2002). Okrem toho elektrofyziologické štúdie na makakoch (Pons a kol., 1987) a údaje z magnetoencefalografie získané u ľudí naznačujú, že sériové spracovanie hmatových vstupov prebieha od S1 po S2 u vyšších primátov (Frot a Mauguiere 1999; Disbrow a kol. ., 2001). Niekoľko štúdií preukázalo, že BA2 je prednostne aktivovaný tvarom a zakrivením (Bodegard a kol., 2001), zatiaľ čo S2 sa môže podieľať na hmatovom učení (Ridley a Ettlinger 1976; Murray a Mishkin, 1984), čo podporuje názor, že tieto oblasti mozgu sa podieľajú na somatosenzorickom spracovaní vyššieho rádu. Naše psychofyzikálne hodnotenie preukázalo, že pacienti boli významne oslabení pri detekcii jednoduchých hmatových stimulov a v teste identifikácie textúry tvaru 1.5 roka po operácii (Taylor et al., 2008a). Tento posledný test hodnotí schopnosť pacienta rozpoznať vlastnosti objektu pri aktívnom skúmaní tvaru alebo štruktúry, čo si vyžaduje integráciu zmyslových informácií v rôznych oblastiach tela (Rosen a Lundborg, 1998). Celkovo možno povedať, že jednou z interpretácií našich údajov je, že pacienti s PNIr sa viac venujú vibrotaktilným stimulom, čo vedie k zvýšenej aktivácii siete pozitívnej úlohy a BA1/3. U týchto pacientov však naše údaje naznačujú, že oblasti spracovania vyššieho rádu, ako napríklad BA2 a S2, nedostali hmatové informácie, čo zase môže viesť k stenčeniu kôry a zníženiu BOLD odpovedí.
Predpokladá sa, že ostrovček hrá úlohu pri integrácii multimodálnych informácií dôležitých pre senzomotorické, emocionálne, alostatické/homeostatické a kognitívne funkcie (Devinsky a kol., 1995; Critchley, 2004; Craig, 2008) a bol označený za limbický senzorický kortex (Craig , 2008). Niekoľko štúdií uvádza aktiváciu ostrovčeka ako odpoveď na hmatovú stimuláciu (Gelnar et al., 1998;
Downar et al., 2002) a štúdie anatomického sledovania u primátov preukázali, že ostrovček je recipročne spojený s frontálnymi, parietálnymi a temporálnymi lalokmi (Augustine, 1996). U našich pacientov bola pravá predná insula jedinou kortikálnou oblasťou, ktorá vykazovala významné kortikálne stenčenie v spojení so zníženými hodnotami frakčnej anizotropie v susednej bielej hmote, čo naznačuje, že kortikálne stenčenie v tejto oblasti je spojené so stratou vlákien vyčnievajúcich do alebo z túto štruktúru. Pravá predná insula sa podieľa na interocepcii, pretože je situovaná tak, aby integrovala homeostatický vstup z tela s motivačnými, emocionálnymi a sociálnymi podmienkami (Craig, 2008). Okrem toho Critchley a kol. (2004) uviedli koreláciu medzi interoceptívnymi schopnosťami a objemom šedej hmoty pravej prednej insuly. Vzhľadom na naše zistenie, že pacienti majú zníženú šedú hmotu v pravej prednej insule, bolo by zaujímavé posúdiť interoceptívne schopnosti po poranení periférnych nervov v budúcej štúdii.
Celkovo sme prvýkrát preukázali, že funkčné a štrukturálne zmeny sú prítomné v ľudskej mozgovej kôre 1.5 roka po kompletnej transekcii periférneho nervu hornej končatiny, ktorý bol mikrochirurgicky opravený. Okrem toho merania nervového vedenia naznačujú u týchto pacientov neúplnú periférnu regeneráciu. Ďalej sme ukázali, že hrúbka kôry súvisí s psychofyzikálnymi mierami zotavenia, v tom, že tenšia kôra v rámci BA2 a S2 bola spojená s horšou somatosenzorickou funkciou. Tieto údaje naznačujú, že obnovenie normálnych funkčných aktivačných máp je priamo spojené s úspešnou regeneráciou periférnych aferentov.
Keri S. Taylor,1,2 Dimitri J. Anastakis2,3,4 a Karen D. Davis1,2,3
1 Division of Brain, Imaging and Behavior � Systems Neuroscience, Toronto Western Research Institute, University Health Network, Toronto, Kanada M5T258
2 Inštitút lekárskej vedy, University of Toronto, Kanada
3 Katedra chirurgie, University of Toronto, Kanada
4 Centrum zdrojov pre klinické štúdie, Toronto Western Research Institute, University Health Network, Toronto, Kanada M5T2S8
Korešpondencia s: Karen D. Davis, Ph.D.,
Division of Brain, Imaging and Behavior � Systems Neuroscience, Toronto Western Research Institute,
Západná nemocnica v Toronte,
Univerzitná zdravotnícka sieť,
Izba MP14-306, 399 Bathurst Street,
Toronto, Ontário,
Kanada M5T 2S8
E-mail: kdavis@uhnres.utoronto.ca
Poďakovanie
Autori ďakujú pánom Geoffovi Popeovi, Dr. Adrianovi Crawleymu, Eugenovi Hlasnému a Keithovi Taovi za odbornú technickú pomoc. Autori by sa chceli poďakovať Dr. Petrovi Ashbymu a pánovi Freddymu Paizovi z EMG kliniky Západnej nemocnice v Toronte za vykonanie testov nervového vedenia a za poskytnutie odborného posúdenia zistení. Autori tiež ďakujú Drs Dvali, Binhammer, Fialkov a Antonyshyn za spoluprácu na tomto projekte. Dr. Davis je kanadským vedúcim výskumu v oblasti mozgu a správania (CIHR MOP 53304).
Financovanie
The Physicians� Services Incorporated a spoločný počiatočný grant od University of Toronto Center for the Study of Pain/AstraZeneca.
Doplnkový materiál
Doplnkový materiál je dostupný na Brain online.
prázdny
Referencie:
Apkarian AV, Sosa Y, Sonty S, a kol. Chronická bolesť chrbta je spojená s
znížená hustota prefrontálnej a talamickej šedej hmoty. J Neurosci
2004; 24: 10410�5.
Arthurs OJ, Johansen-Berg H, Matthews PM, Boniface SJ. Pozornosť
diferenciálne moduluje spojenie fMRI BOLD a evokované
amplitúdy potenciálnych signálov v ľudskom somatosenzorickom kortexe 7.
Exp Brain Res 2004; 157: 269.
Augustín JR. Obvodové a funkčné aspekty ostrovného laloku
u primátov vrátane ľudí. Brain Res Brain Res Rev 1996; 22:
229.
Beaulieu C. Základ anizotropnej difúzie vody v nervovom
systém – technický prehľad. NMR Biomed 2002; 15: 435.
Bodegard A, Geyer S, Grefkes C, Zilles K, Roland PE. Hierarchický
spracovanie hmatového tvaru v ľudskom mozgu. Neuron 2001; 31:
317.
Craig AD. Interocepcia a emócie: Neuroanatomická perspektíva.
In: Lewis M, Haviland-Jones J, Barrett L, redaktori. Príručka o
emócie. New York: Guildford Press; 2008. s. 272.
Critchley HD. Ľudská kôra reaguje na interoceptívnu výzvu.
Proc Natl Acad Sci USA 2004; 101: 6333�4.
Critchley HD, Wiens S, Rotshtein P, Ohman A, Dolan RJ. Neurónové systémy
podpora interoceptívneho povedomia. Nat Neurosci 2004; 7: 189.
Dale AM, Fischl B, Sereno MI. Kortikálna povrchová analýza. I. Segmentácia
a rekonštrukcia povrchu. Neuroimage 1999; 9: 179.
Davis KD, Pope G, Chen J, Kwan CL, Crawley AP, Diamant NE. Kortikálna
rednutie pri IBS: dôsledky pre homeostatiku, pozornosť a bolesť
spracovanie. Neurológia 2008; 70: 153.
DeLuca M, Beckmann CF, De SN, Matthews PM, Smith SM. fMRI v pokoji
štátne siete definujú odlišné režimy interakcií na veľké vzdialenosti
v ľudskom mozgu. Neuroimage 2006; 29: 1359-67.
Devínsky O, Morrell MJ, Vogt BA. Príspevky predného cingulátu
kôry k správaniu. Brain 1995; 118 (Pt 1): 279�306.
Disbrow E, Roberts T, Poeppel D, Krubitzer L. Dôkazy pre interhemisférické
spracovanie vstupov z rúk v ľudskom S2
a PV. J Neurophysiol 2001; 85: 2236-44.
Downar J, Crawley AP, Mikulis DJ, Davis KD. Citlivá kortikálna sieť
stimulovať nápadnosť v neutrálnom behaviorálnom kontexte naprieč viacerými
zmyslové modality. J Neurophysiology 2002; 87: 615-20.
Draganski B, Moser T, Lummel N, a kol. Zníženie šedej talamu
záležitosť po amputácii končatiny. Neuroimage 2006; 31: 951�7.
Fischl B, Dale AM. Meranie hrúbky ľudskej mozgovej kôry
zo snímok magnetickej rezonancie. Proc Natl Acad Sci USA 2000; 97:
11050.
Fischl B, Sereno MI, Dale AM. Kortikálna povrchová analýza. II:
Nafukovanie, sploštenie a povrchový súradnicový systém.
Neuroimage 1999a; 9: 195-207.
Fischl B, Sereno MI, Tootell RB, Dale AM. Medzipredmet s vysokým rozlíšením
spriemerovanie a súradnicový systém pre povrch kôry. Hum mozog
Mapp 1999b; 8: 272.
Florence SL, Kaas JH. Rozsiahla reorganizácia na viacerých úrovniach
somatosenzorická dráha nasleduje po terapeutickej amputácii ruky
u opíc. J Neurosci 1995; 15: 8083�95.
Fox MD, Snyder AZ, Vincent JL, Corbetta M, Van E, Raichle ME. The
ľudský mozog je vnútorne organizovaný do dynamických, antikorelovaných
funkčné siete. Proc Natl Acad Sci USA 2005; 102: 9673-9678.
Friedman DP, Murray EA. Thalamická konektivita druhého
somatosenzorická oblasť a susedné somatosenzorické polia
laterálny sulcus makaka. J Comp Neurol 1986; 252: 348.
Frot M, Mauguiere F. Časovanie a priestorové rozloženie somatosenzorických funkcií
odozvy zaznamenané v hornom brehu sylvianskej pukliny (oblasť SII) v
ľudí. Cereb Cortex 1999; 9: 854-63.
Geha PY, Baliki MN, Harden RN, Bauer WR, Parrish TB, Apkarian AV.
Mozog pri chronickej bolesti CRPS: abnormálna šedo-biela hmota
interakcie v emocionálnych a autonómnych oblastiach. Neuron 2008; 60:
570.
Gelnar PA, Krauss BR, Szeverenyi NM, Apkarian AV. Znázornenie končekov prstov
v ľudskej somatosenzorickej kôre: štúdia fMRI.
Neuroimage 1998; 7: 261.
Hickmott PW, Steen PA. Rozsiahle zmeny v dendritickej štruktúre
počas reorganizácie somatosenzorickej kôry dospelých. Nat Neurosci
2005; 8: 140�42.
Iwamura Y, Tanaka M, Iriki A, Taoka M, Toda T. Spracovanie
taktilné a kinestetické signály z bilaterálnych strán tela v
postcentrálny gyrus bdelých opíc. Behav Brain Res 2002; 135:
185.
Jaquet JB, Luijsterburg AJ, Kalmijn S, Kuypers PD, Hofman A, Hovius SE.
Stredné, ulnárne a kombinované poranenia stredno-ulnárneho nervu: funkčné
a návrat k produktivite. J Trauma 2001; 51: 687.
Jurkiewicz MT, Crawley AP, Verrier MC, Fehlings MG, Mikulis DJ.
Somatosenzorická kortikálna atrofia po poranení miechy: založená na voxeli
morfometrická štúdia. Neurológia 2006; 66: 762�4.
Kaas JH. Plasticita senzorických a motorických máp u dospelých cicavcov. Annu
Rev Neurosci 1991; 14: 137.
Kaas JH, Jain N, Qi HX. Organizácia somatosenzorického systému v
primátov. In: Nelson RJ, redaktor. Somatosenzorický systém.
Washington, DC: CRC Press; 2002. s. 1�25.
Kimura J. Elektrodiagnostika pri ochoreniach nervov a svalov: princípy
a prax. Oxford: Oxford University Press; 2001.
Kimura J. Princípy a úskalia štúdií nervového vedenia. Ann Neurol
1984; 16: 415�29.
Knyihar-Csillik E, Rakic P, Csillik B. Transneuronálna degenerácia v
látka Rolando miechy primátov vyvolaná tým
axotómiou indukovaná transgangliová degeneratívna atrofia centrál
primárne senzorické terminály. Cell Tissue Res 1989; 258: 515.
Liss AG, af Ekenstam FW, Wiberg M. Strata neurónov v dorzálnom koreni
gangliá po prerezaní periférneho senzorického nervu. Anatomický
štúdium na opiciach. Scand J Plast Reconstr Surg Surg ruky 1996; 30: 1�6.
Lotze M, Flor H, Grodd W, Larbig W, Birbaumer N. Fantómové pohyby
a bolesť. Štúdia fMRI u pacientov s amputáciou hornej končatiny. Brain 2001;
124: 2268-77.
Manduch M, Bezuhly M, Anastakis DJ, Crawley AP, Mikulis DJ. Serial
fMRI adaptačných zmien v primárnej senzomotorickej kôre
rekonštrukcia palca. Neurológia 2002; 59: 1278-81.
Máj A. Chronická bolesť môže zmeniť štruktúru mozgu. Bolesť 2008;
137: 7-15.
McAllister RM, Gilbert SE, Calder JS, Smith PJ. Epidemiológia a
manažment poranení periférnych nervov hornej končatiny v modern
prax. J Hand Surg (Br) 1996; 21: 4:13.
Murray EA, Mishkin M. Relatívne príspevky SII a oblasti5 k hmatu
diskriminácia u opíc 2. Behav Brain Res 1984; 11: 67-83.
Napadow V, Kettner N, Ryan A, Kwong KK, Audette J, Hui KK.
Somatosenzorická kortikálna plasticita pri syndróme karpálneho tunela - prierez
hodnotenie fMRI. Neuroimage 2006; 31: 520-30.
Oldfield RC. Hodnotenie a analýza ruky: Edinburgh
inventár. Neuropsychologia 1971; 9: 97.
Pons TP, Garraghty PE, Cusick CG, Kaas JH. Somatotopická organizácia
oblasti 2 u makakov 6. J Comp Neurol 1985; 241: 445-66.
Pons TP, Garraghty PE, Friedman DP, Mishkin M. Fyziologické dôkazy
na sériové spracovanie v somatosenzorickom kortexe. Science 1987; 237:
417.
Porro CA, Lui F, Facchin P, Maieron M, Baraldi P. Aktivita súvisiaca s vnímaním
v somatosenzorickom systéme človeka: funkčná magnetická rezonancia
zobrazovacie štúdie. Magn Reson Imaging 2004; 22: 1539-48.
Powell TP, Erulkar S. Degenerácia transneuronálnych buniek v sluchovej oblasti
reléové jadrá kat. J Anat 1962; 96: 249-68.
Ridley RM, Ettlinger G. Zhoršené hmatové učenie a uchovanie po
odstránenie druhej somatickej senzorickej projekčnej kôry (SII) v
opice. Brain Res 1976; 109: 656.
Rosen B, Lundborg G. Nový nástroj taktilnej gnózy v citlivosti
testovanie. J Hand Ther 1998; 11: 251�7.
Seminowicz DA, Davis KD. Bolesť zvyšuje funkčnú konektivitu
mozgovej siete vyvolanej výkonom kognitívnej úlohy.
J Neurophysiol 2007; 97: 3651�9.
Smith SM, Jenkinson M, Johansen-Berg H a kol. Traktový priestorový
štatistika: voxelová analýza údajov o difúzii viacerých subjektov.
Neuroimage 2006; 31: 1487.
Talairach J, Tournoux P. Koplanárny stereotaxický atlas ľudského mozgu.
New York: Thieme Medical Publishers Inc.; 1988.
Taylor KS, Anastakis DJ, Davis KD. Chronická bolesť po periférnej
poranenie nervov je spojené s katastrofickou bolesťou a neurotizmom.
Int Ass Stud Pain 2008a; 267.
Taylor KS, Davis KD. Stabilita fMRI súvisiacej s hmatom a bolesťou
aktivácie mozgu: vyšetrenie prahovo závislých a
prahovo nezávislé metódy. Hum Brain Mapp 2009; 30:
1947.
Taylor KS, Seminowicz DA, Davis KD. Dva systémy pokojového stavu
konektivitu medzi ostrovčekom a cingulárnou kôrou. Hum mozog
Mapa 2008b;DOI:10.1002/hbm.20705.
Wall JT, Kaas JH, Sur M, Nelson RJ, Felleman DJ, Merzenich MM.
Funkčná reorganizácia v somatosenzorických kortikálnych oblastiach 3b a 1
dospelých opíc po oprave stredného nervu: možný vzťah k
regenerácia zmyslov u ľudí. J Neurosci 1986; 6: 218.
Zatvorte akordeón
Informácie tu uvedené o „Podrezanie nervov zmení váš mozog | El Paso, TX." nie je určený na nahradenie vzťahu jeden na jedného s kvalifikovaným zdravotníckym pracovníkom alebo licencovaným lekárom a nie je to lekárska rada. Odporúčame vám, aby ste rozhodnutia v oblasti zdravotnej starostlivosti robili na základe vášho výskumu a partnerstva s kvalifikovaným zdravotníckym pracovníkom.
Informácie o blogu a diskusie o rozsahu
Náš informačný rozsah sa obmedzuje na chiropraktické, muskuloskeletálne, fyzické lieky, wellness, prispievajúce etiologické viscerozomatické poruchy v rámci klinických prezentácií, súvisiacej somatoviscerálnej reflexnej klinickej dynamiky, subluxačných komplexov, citlivých zdravotných problémov a/alebo článkov, tém a diskusií o funkčnej medicíne.
Poskytujeme a prezentujeme klinická spolupráca so špecialistami z rôznych odborov. Každý špecialista sa riadi svojím odborným rozsahom praxe a jurisdikciou udeľovania licencií. Funkčné zdravotné a wellness protokoly používame na liečbu a podporu starostlivosti o zranenia alebo poruchy pohybového aparátu.
Naše videá, príspevky, témy, predmety a postrehy pokrývajú klinické záležitosti, problémy a témy, ktoré sa týkajú a priamo či nepriamo podporujú náš klinický rozsah praxe.*
Naša kancelária sa primerane pokúsila poskytnúť podporné citácie a identifikovala relevantnú výskumnú štúdiu alebo štúdie podporujúce naše príspevky. Na požiadanie poskytujeme kópie podporných výskumných štúdií, ktoré majú regulačné rady a verejnosť k dispozícii.
Rozumieme, že pokrývame záležitosti, ktoré si vyžadujú ďalšie vysvetlenie, ako môže pomôcť v konkrétnom pláne starostlivosti alebo v protokole liečby; na ďalšiu diskusiu o vyššie uvedenej téme sa preto môžete pokojne opýtať Dr. Alex Jimenez, DC, Alebo kontaktujte nás na adrese 915-850-0900.
Sme tu, aby sme vám a vašej rodine pomohli.
Požehnanie
Dr. Alex Jimenez DC MSACP, RN*, CCST, IFMCP*, CIFM*, ATN*
e-mail: coach@elpasofunctionalmedicine.com
Licencovaný ako doktor chiropraxe (DC) v Texas & Nové Mexiko*
Číslo licencie Texas DC TX5807, New Mexico DC Licencia č. NM-DC2182
Licencovaná ako registrovaná zdravotná sestra (RN*) in Florida
Floridská licencia RN licencia # RN9617241 (Kontrola č. 3558029)
Kompaktný stav: Viacštátna licencia: Oprávnený vykonávať prax v Štáty 40*
Dr. Alex Jimenez DC, MSACP, RN* CIFM*, IFMCP*, ATN*, CCST
Moja digitálna vizitka





